
1. Introduction
Chromium (Cr) is one of the environmental pollutants originated from various industrial wastes. For instance, chromium (VI) and chromium (III) are used in the plating, textile, leather tanning, and wood preservation industries [1]. In general, chromium has two main forms in the soil
environment [2-3], i.e., the dominant form chromium (III) that is under a reduced conditions, immovable, and less toxic to plants, and the oxidized form chromium (VI) that is very easy to move and carcinogenic to humans [ 3]. Changes in the concentration of Cr (VI) on soil can affect its absorption by plants, caused inhibited growth, disruption of photosynthesis and respiration, absorption of 1) School of Life Sciences and Technology, Bandung Institute of Technology, Indonesia.
minerals, enzyme activity, and ended with damage to membrane lipids and plant DNA [4].
Water hyacinth including the Pontederiaceae family is found in the tropics and subtropics. Water hyacinths are classified as aquatic weeds that can adapt to changes in the environment and multiply rapidly. The ideal growing place for water hyacinth plants is shallow and turbid waters, with temperatures ranging from 28-30 °C and pH conditions ranging from 4-12. In deep and clear waters in the highlands, these plants are difficult to grow. Water hyacinth can suck water and vaporize it into air water through the transpiration process [5].
Pistia stratiotes are sometimes used as decorative plants in a pond and play an important role for phytoremediation of heavy metals, chemical products and radioactive material in water [6]. Different accumulations of Cr in parts of the plant have been reported. The root is the main organ in plants that is directly related to the absorption of nutrients from the environment, including heavy metal like Cr, Pb, Ni, etc. In beans, at least 0.1% of the total Cr that accumulates was found in the seeds, whereas the concentration in the roots reached 98% of the total absorption of Cr [7]. The amount of Cr accumulations (160- 350 mg Cr / kg of dry weight at the root and reaching 1.6-2.0 mg Cr / kg dry weight on shoots) was observed for cauliflower, green vegetables, and chili. A high capacity to accumulate Cr was shown by water hyacinth (Eichhornia crassipes) with up to 6 mg/ day dry weight in the roots of this plant that grows at 10 ppm Cr [VI].
A study reported the toxicity of Cr on plant growth and development (i.e., change in germination process followed by changes in root, stem and leaf growth) can affect the total production of plant dry weight [8]. The inhibition of chlorophyll biosynthesis has also been reported in land plants [8-9]. For example, barley seedlings grown at 100 mM Cr showed a 40% lower growth rate [8]. Chromium also damaged the plants' physiological processes, i.e., photosynthesis, absorption of water, and nutrients absorption. Metabolic changes also occur with the production of enzymes and metabolites or by producing Reactive Oxygen Species (ROS) [10].
Heavy metals can have an ecological impact on water bodies leading to increased nutrient load especially if they are essential metals. These metals in the effluent may increase the fertility of water leading to eutrophication which in open water can progressively lead to oxygen deficiency, algae blooms, and death of aquatic life [11].
Chlorophyll and proline are some examples of plant metabolism antioxidant products [12] which play a role in the major metal detoxification mechanisms in plants [13]. Environmental changes such as pollution from industrial waste will alternate the physiological responses of the plant. The implication of this study is to study the changes in physiological plants as a trivalent chromium bioaccumulator agent.
2. Materials and methods
2.1. Plant material and experimental design
Water hyacinth and water lettuce with almost the same size for each plant (based on weight) was collected from a pond in ITB campus and with an analytical grade of Cr3+ (CrCl3). The experimental design consists of four treatments, i.e., control, Cr3+ 40 ppm, 80 ppm, and 120 ppm.
There were three replicates for each treatment. Twelve plants of each were placed in 10 L of medium. The experiment was conducted for 2 weeks at the screen house. After two weeks, the plants were harvested and weight was measured. The samples were separated into shoot and root and were analyzed for Cr content, chlorophyll, proline and CAT enzyme activity.
2.2. Cr content in the media, roots, and leaves
Cr content in the leaves and roots was measured by concentrating the sample with nitric acid (HNO3) [14]. The accumulation of Cr in Eichhornia crassipes and Pistia stratiotes was determined by calculating the concentration of chromium in its water, roots, and leaves. A comparison between the root/leaves concentration and concentration in water is known as Bio-Concentration Factor (BCF). BCF in leaves and roots was calculated to determine how much metal concentration in leaves and roots was assumed to be from the environment [15]. The formula used to calculate BCF was as follows:
\[BCF = \frac{\text{Cr in roots or leaves (mg/Kg)}}{\text{Cr in media (mg/Kg)}}\](1)
A comparison between the metal concentrations in leaves and roots known as Translocation Factors (TF) was also calculated. TF values were calculated to determine the displacement of metal accumulation from root to shoot [16]. TF values were calculated using the formula:
\[TF = \frac{\text{Cr in leaves (mg/Kg)}}{\text{Cr in roots (mg/Kg)}}\](2)
The differences between BCF and TF values were then used to calculate Phytoremediation (FTD) [16]. FTD was calculated using the formula:
\[FTD = BCF - TF \tag{3}\]
2.3. Chlorophyll content
Measurement of chlorophyll levels was carried out based on [17]. Fresh leaf samples were weighed 0.1 grams and then crushed using 80% acetone. The chlorophyll solution was then filtered into chlorophyll filtrate and then taken for absorbance measurements using a spectrophotometer at a wavelength of 645 nm and 663 nm. Chlorophyll levels were calculated using the formula:
Total chlorophyll (mg/g) = \[\frac{8,02 A663+20,2 A645}{1000 FW}\] x V (4)
V = sample volume A = absorbance value FW = sample fresh weight (g)
2.4. Proline levels
Proline level measurement was carried out by measuring the absorbance using a spectrophotometer at a wavelength of 520 nm [18]. Proline levels were calculated based on the proline standard curve based on fresh weight using the formula:
\[\mu \text{mol Prolin/g FW} = \frac{\mu \text{g Prolin x mL toluene}}{115.5} x \frac{5}{\text{g sample}}\](5)
115.5 = proline molecule weight
2.5. Catalase enzyme activity (CAT)
CAT enzyme activity was measured based on the method carried out by [19]. CAT enzyme activity was measured using a spectrophotometer by measuring changes in sample absorbance at a wavelength of 240 nm as a consequence of H2O2 degradation. Readings of absorbance numbers were done every 30 seconds for three minutes. The activity of catalase enzyme was calculated using the formula:
Volume activity (unit/mL) = \[\frac{\Delta A \, x \, Vq}{0.0436 \, x \, Vs}\] (6)
CAT enzyme activity (unit/mg) = \[\frac{\text{volume activity}}{\text{fresh weight}}\] (7)
\(\Delta A = Absorbance\) at third minute - Absorbance at initial minute
Vq = Volume reaction in kuvet (ml)
Vq = Volume sample (ml)
2.6. Statistical analysis
Data were statistically analyzed using a statistic software IBM SPSS Statistics version 25. One-way ANOVA was used for proline, catalase enzyme, and chlorophyll content analysis. The results were presented in graphs. Data from different treatments and control were compared by Duncan's multiple range test (DMRT) at p < 0.05.
3. Results
3.1. Cr Level on Roots, Leaves and Medium
According to the result (Figure 1 and Figure 2), the Cr content in the roots of water lettuce and water hyacinth has increased from control to the concentration of 80 ppm, but at the concentration of 120 ppm, it has decreased.
Figure 3 showed the ability of water hyacinth and water lettuce as a phytoremediation agent. The highest FTD value obtained from water lettuce root at Cr level 40 ppm, 80 ppm, and 120 ppm.
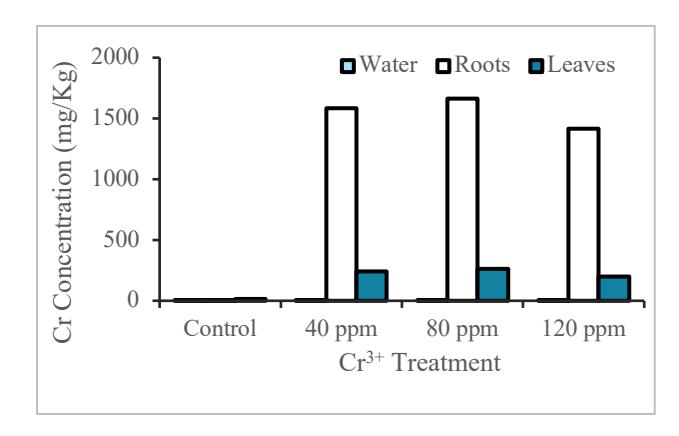
Figure 1 Analysis of Cr in water lettuce
3.2. Chlorophyll Level
Figure 4 showed that at concentrations of 0 to 80 ppm, the chlorophyll content in water hyacinth is above the chlorophyll content of water lettuce, while at the concentration of 120 ppm, the chlorophyll content of water lettuce is slightly higher. The statistical test showed no significant difference between the chlorophyll content of the two plants for each different Cr concentration (P>0.05).
3.3. Catalase Enzyme Activity
Both water lettuce and water hyacinth plants showed decrease in CAT enzyme activity along with the increase in Cr metal in water media (Figure 6).
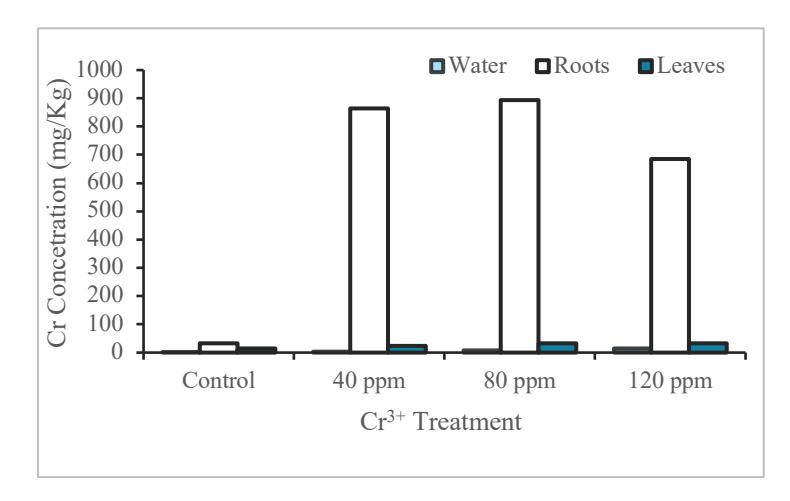
Figure 2 Analysis of Cr in water hyacinth
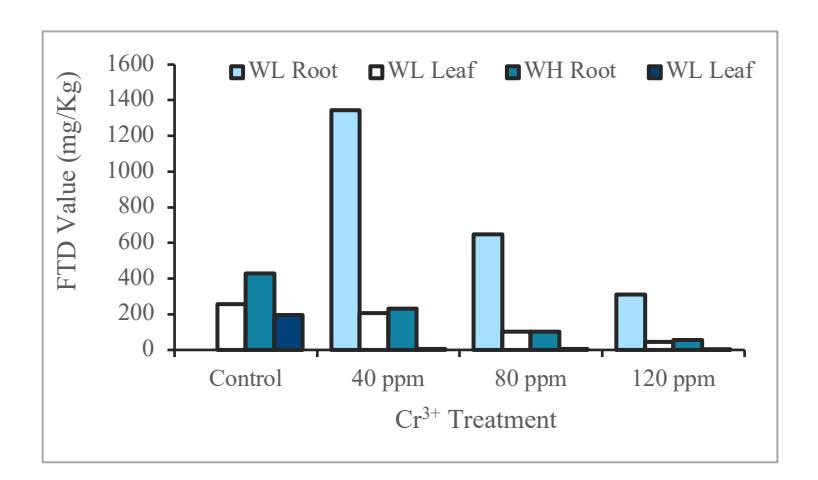
Figure 3 Graph of FTD value in water hyacinth and water lettuce
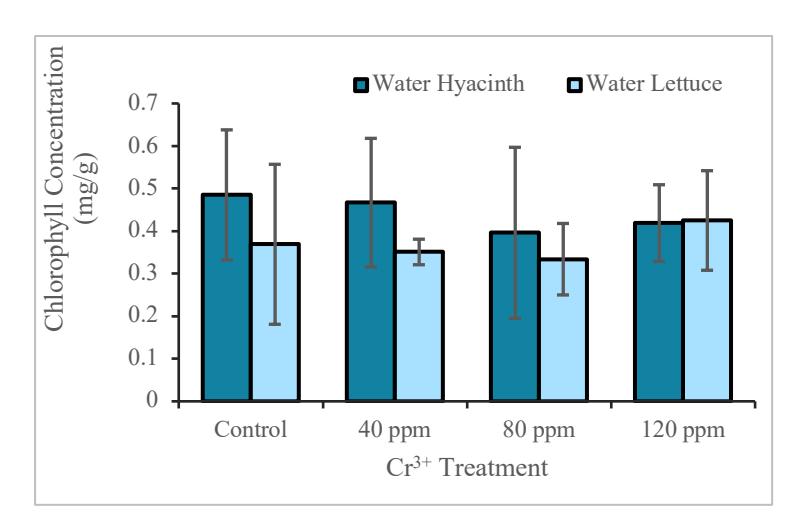
Figure 4. Graph of chlorophyll level in water hyacinth and water lettuce
Figure 5 Comparison of water hyacinth in control treatment and 80 ppm treatment
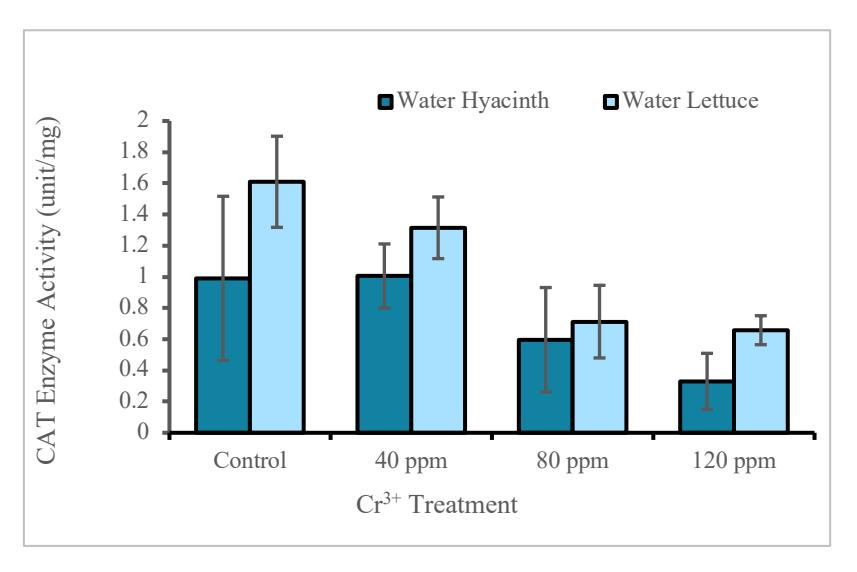
Figure 6. Graph of CAT enzyme activity in water hyacinth and water lettuce
3.4. Plant Biomass
In water hyacinth plants, the sample with 0 ppm treatment (control), has the biggest difference value between initial and final fresh weight (Table 1) compared to the water hyacinth treated by Cr. In water hyacinth, Cr can inhibit vegetative growth. There was no initial dry weight measurement to keep the plants alive for three weeks (time of treatment).
4. Discussion
The results of the analysis of Cr3+ in water showed that the water hyacinth plant samples have a higher Cr content compared with water lettuce plant. The low Cr3+ content in ppm for water lettuce plants, but still, there is no evidence for this mechanism especially for Cr3+ absorption.
water lettuce water samples showed that the absorption of Cr3+ by water lettuce is more effective than water hyacinth. Cr3+ treatment of water lettuce showed growth differences that produce a lot of small vegetative plants (tillers). Another research showed that P. stratiotes was able to efficiently reduce Cr6+ in 16 days, thus this plant has a good performance in reducing Cr6+ [20].
As shown in Figure 1 and 2, Cr3+ content on the roots of water lettuce is higher than water hyacinth. For both water hyacinth and water lettuce, Cr3+ content from the control treatment up to 80 ppm was increased but started to decrease at 120 ppm concentration. This shows the maximum ability of Cr absorption was likely to reach a concentration of 80
Table 1. Plant biomass on different treatments
| Weight (gram) | Water Hyacinth | Water Lettuce | ||||||
|---|---|---|---|---|---|---|---|---|
| Control | 40 ppm | 80 ppm | 120 ppm | Control | 40 ppm | 80 ppm | 120 ppm | |
| Initial Weight | 170 | 225 | 207 | 188 | 174 | 143 | 113 | 106 |
| Final Weight | 410 | 421 | 427 | 302 | 231 | 274 | 182 | 255 |
| Dry weight | 15.38 | 13.14 | 12.14 | 4.62 | 8.48 | 8.77 | 5.20 | 7.43 |
A previous study on P. stratiotes treated with Cr6+ showed that the plant could effectively remove Cr6+ from the solution with minor damage up to an initial Cr6+ concentration of 3 mg/L, for which the adsorption isotherm studies were conducted. The maximum uptake capacity of the plant was recorded was 0.05 mg/g of Cr (VI) at the equilibrium level after a contact period of 7 days for an initial concentration of 8 mg /L, although severe physiological damage occurred [21].
The results of Cr analysis on the leaves of water lettuce are higher than those of water hyacinth. In the analysis of water hyacinth leaves, the higher Cr concentration coincides with the higher the Cr leaf content. However, on water lettuce, the control treatment up to 80 ppm experienced an increase of Cr concentration and at a concentration of 120 ppm, it decreased. In Figure 1 and 2, the concentration of Cr in water lettuce is higher than water hyacinth, although, at the concentration of 0 ppm the Cr content is slightly higher in water hyacinth, it is related to the mobility of heavy metals in plants. Each species has its own mechanism for stress, including heavy metal stress. Plasma membranes act as a barrier for heavy metal ions to be able to enter cells, so this plasma membrane plays an important position in plants that are resistant to heavy metals [22]. In water lettuce, although the absorption and mobility of Cr is high in the body, it affected the growth and increased mortality rate. Water hyacinth, however, can last longer in Cr contaminated water because the levels of Cr absorbed and the mobility of Cr are low in the body.
Based on this, water lettuce can be used for phytoremediation purposes in Cr polluted environments. Phytoremediation is one solution that is inexpensive, sustainable, and saves energy in contaminated areas. Phytoremediation is the application of plants for in-situ or ex-situ treatment/removal of contaminated soils, sediments and water [23]. One example of phytoremediation is phytostabilization [24]. Phytostabilization is an attempt to reduce the pollutant content in which plants are used as an agent to reduce heavy metal movement in the soil. Based on the results of the Cr test, the higher increasing of Cr level in the water, the higher the level Cr in plant tissue. The Cr plant tissue increased along with the increase of Cr concentration in water. This is because the concentration of Cr solution is more concentrated than in the root tissue, so Cr enters the root [25].
According to research in [25], the roots accumulate 10- 100 times more than the shoots and other tissues. Several studies show that in general, the absorption capacity of heavy metals in plants followed a sequence: roots, stems, and leaves [26]. Cr toxic effects can be seen from plant morphology. The leaves became yellow, some almost come off the trunk, and some fall out. Other toxic effects also occur in the roots where some of the root fibers are separated from the roots. Cr in high concentrations also inhibits enzyme activity, damages root cells, and causes ultrastructural modifications to chloroplast and cell membranes [27]. This is consistent with the observations shown in Figure 5.
The Cr absorption is higher in water lettuce than water hyacinth. The difference in absorption is influenced by the characteristics of water plants. Water hyacinth can bind organic materials from mud particles which make this plant suitable to purify water. This plant also serves an ecological function as the water stabilizer due to its ability to neutralize pollutants in the waters [28]. Through its thick roots, Cr metal is absorbed and then used in metabolic processes or stored in roots, stems, tubers or leaves; and can absorb excess nutrients in the water. Water lettuce is very resistant to the low nutrient levels in the water but its response as well to high nutrient levels by forming a lot of vegetative tillers [28]. This is what causes the water lettuce to be more resistant compared to water hyacinth. Water hyacinth could absorb Cr as well as water lettuce but could not form a lot of tillers. Cr affect water uptake of plants by disrupting growth and development including changes in the germination process followed by changes in root, stem and leaf growth.
Several studies reported the plants' response to stresses that affect chlorophyll levels, such as drought stress. According to [29], chlorophyll concentration can be used as an indicator of the lack of water in plants. Figure 4 shows that at concentrations of 0 to 80 ppm the chlorophyll content in water hyacinth was above the chlorophyll content of water lettuce; while at 120 ppm Cr concentration, the chlorophyll content of water lettuce was slightly higher. From the statistical test, it was found that there were no significant differences between the chlorophyll levels of the two plants for each different Cr concentration (P> 0.05). Heavy metals affect photosynthesis by influencing pigment formation, electron transport activity, and calvin cycle enzymes. Cr plays a role in inhibiting thylakoid formation in chloroplasts
[30]. Besides, the main target of heavy metals is in the electron transport process. Although there was no significant difference in water hyacinth which was treated with the addition of Cr, chlorosis can be seen morphologically, i.e., a condition in which plant tissue fails to form chloroplasts resulting in pale green or yellowish leaf color. In this study the differences seen from the color of water hyacinth leaves as shown in Figure 5. Figure 5 showed the chlorosis in leaves of water hyacinth and for water lettuce the chlorosis was not seen.
This research showed that there was a decrease in proline levels in water hyacinth plants along with the increase in the concentration of Cr; however, there were no statistically significant differences. A research by [31] on the effect of Cr on photosynthetic pigments, stress defense systems, nitrate reduction and proline levels on Ocinum tenuiflorum plants showed that the higher the concentration of Cr, the higher the level of proline, which was thought to be a form of defense against heavy metal stress. In this study, there was a decrease in proline levels with the increase in Cr, which can occur due to the administration of Cr3+ with low toxicity [3], therefore, water hyacinth plants do not respond to Cr3+ as a metal compound that causes stress.
In contrast to the response of water hyacinth, the proline levels in water lettuce showed that a low proline leveil in control and 40 ppm. However, the proline levels were quite high at 80 ppm concentration decreased at a concentration of 120 ppm. Several studies on plants responses to the changes in water content (drought, salinity, freezing) showed the formation of proline was one way to defend the plant [32]. Concerning heavy metal stress, Cr in high concentrations interferes with the process of absorption of water and nutrients by roots, so plants are not able to absorb water optimally and a process occurs that causes the plant to dry out and in response produces proline compounds.
The activity of catalase (CAT) enzyme in water hyacinth did not differed significantly (p> 0.05) in each treatment. For water lettuce, there was a significant difference (p <0.05) on the addition of CrCl3 between a concentration of 80 ppm and 120 ppm with the control. Based on Figure 5, both water lettuce and water hyacinth experienced a decrease in CAT along with the increase in Cr (III) metal in water media. This is different from the results obtained by [33] who observed an increasing CAT with variations in Cr metal concentration from 0 to 5 mM. Another study by [34] showed a decreasing catalase enzyme activity in water hyacinth exposed to Cd between 50 and 75 ppm. CAT measurement is one of the groups of antioxidant enzymes that are commonly analyzed in plants exposed to heavy metals. These enzymes are peroxidase (POD) and superoxide dismutase (SOD). These tenzymes have the highest activity on plant roots, because roots are the initial entry of heavy metals into plant tissue systems [35]. The reason CAT decreased along with the increase of Cr3+ was due to the possibility of not forming H2O2. Increase in H2O2 production were reported to occur during a biotic and/or abiotic stress regime, such as pathogen attack, wounding, UV irradiation, exposure to intense light, drought, salinity, and/or chilling [36].
The activity of catalase hyacinth and water lettuce enzymes were shown in Figure 6. Plants can avoid the entry of heavy metal ions into their bodies based on immobilization in the cell wall and then release cell ligands that act as chelating agents for metals. As a result, plants are able to make heavy metals that will enter as a non-toxic substance in intracellular processes [24].
Table 1 showed a change in the amount of biomass in each treatment both for water hyacinth and water lettuce. In water hyacinth, samples from the control treatment have the biggest weight difference between initial fresh weight and final fresh weight (240 gr); and for Cr3+ treatment 120 ppm, the weight difference was 114 gr, which shows that in water hyacinth, Cr3+ may inhibit vegetative growth.
In water lettuce, different results were obtained, the highest difference between initial fresh weight and final fresh weight was obtained from water lettuce with the 120 ppm Cr3+ concentration. Water lettuce is capable of producing vegetative tillers, and the number of vegetative tillers was increased in the treatment of 120 ppm Cr. Although the main plants are not able to live long in high Cr concentrations, but vegetative tillers can absorb Cr by suppressing their body size, resulting in much smaller tillers.
5. Conclusions
Both plants accumulated Cr metal in roots with the largest concentration of 1660 mg / Kg in P. stratiotes and 894.126 mg /Kg in E. crassipes, respectively, with both at 80 ppm treatment. It was found that there was no significant difference (p> 0.05) on chlorophyll and proline levels in each treatment for each species of aquatic plant. The activity of catalase enzyme decreased with increasing concentration of Cr3+ in water media, with a significant difference (P <0.05) for P. stratiotes which was exposed to 80 to 120 ppm compared to the control and from this research water lettuce has better performance for trivalent chromium removal in water.