
1. Introduction
Genetic engineering has made substantial progress in recent years. One of these advances was the discovery of several components that could support the introduction of genetic engineering, for instance, promoters. In genetic engineering, especially in gene introduction, the role of the promoter is very important for the success of protein expression. In process of protein expression, the transcription initiation is a very important initial step, since this process is the starting point for attaching RNA polymerase to the DNA sequence before the transcription
process begins [1]. The DNA sequences where the initiation of transcription (attachment of RNA polymerase and transcription factors) begin are called promoter sequences. Without the promoter sequence, the transcription process will not be induced [2]. The promoter sequence consists of a specific base sequence and serves to support the initiation of transcription [3]. Studies of promoters have been carried out to see the expression patterns of a gene. The control of the promoter can be an important factor that can explain how a gene can be expressed or inhibited.
Studies on promoter have been widely carried out, for instance, on the EF1A gene (Elongation factor 1 alpha).
EF1A presents as abundant proteins in cells, which fills up to 3 - 10% of the total soluble protein in cells [4]. EF1A protein is included in the Moonlight protein group, a protein that has more than one function [5]. This protein can interact with several types of proteins including actin protein [6], microtubules [7], and calmodulin [8]. This protein is also known to play a role in the process of apoptosis [9], regulation of viral infections [10], and DNA replication [11]. Variation in function and high amount of protein makes the EF1A gene family promoter expected to be one source of promoters that can be actively varied and have a high level of expression [12].
Several promoters of the EF1A gene family derived from cassava (Manihot esculenta) have been successfully isolated in previous research [13]. This study isolated promoter of MeEF1A3, MeEF1A5, and MeEF1A6 genes. MeEF1A promoters have different levels of expression in the studied plant model. MeEF1A3 can be expressed in cotyledons but cannot be expressed at the root of tobacco. MeEF1A5 can be expressed in bananas and tomatoes, but cannot be expressed in tobacco shoots. MeEF1A6 can be active in all parts of tobacco shoots, bananas, and tomatoes and has a high level of expression [13]. This makes the MeEF1A6 promoter a constitutive promoter candidate because it could be active continuously and had a very high level of activation. To find out the exact work of the MeEF1A6 promoter, further research is needed on the analysis of this promoter. This study focused on promoter transformation, which was carried out using Agrobacterium tumefaciens strain AGL1 and LBA4404. The results of the promoter expression were carried out by an integrated histochemical GUS assay on the pBI121 plasmid.
2. Material and Methods
2.1. Material
Agrobacterium tumefaciens (AGL1 and LBA4404 Strains) were used as a vector. AGL1 was used from the genetic lab of Bandung Institut of Technology (ITB) and LBA4404 was used from Biorin lab Bogor Agricultural University (IPB). Tobacco plant CV SR1 (Nicotiana tabacum) 3-4 week age was used as a plant model for expression analysis. pBi121 plasmid was used as a plasmid vector. Kanamycin antibiotic was used for plasmid selection, Ampicillin and streptomycin antibiotic was used for selection A tumefaciens strain AGL1 and LBA4404.
2.2. Introduction of MeEF1A6 Promoter to Agrobacterium tumefaciens
Gene introduction was carried out using the electroporation method using electrocompetent cells. Electrocompetent cells were made by following the method
described by Debler [14]. Electrocompeten cells were made by incubating Agrobacterium tumefaciens until it reached exponential phase (2-3 days) and centrifuge gradually until the volume reaches a quarter of the previous volume (every step must conducted in ice-cold temperature). The final pellet was resuspended with 10% glycerol and stored at -80ºC. Introduction plasmid to Agrobacterium tumefaciens begins by mixing 1 μL of the promoted plasmid (plasmid concentration of not more than 20 μg/mL) with 40 μL of electrocompetent cells in the electroporation cuvette. Electroporation conditions were regulated on the Xcell Electroporation System Gene pulser with the choice of Agrobacterium tumefaciens electroporation method. The electroporation cuvette was placed on the Xcell Shock Pod and the electroporation process begins by pressing the pulse button. After the graph appears on the electroporator screen, the culture is quickly transferred to the refresher media (LB medium for LBA4404 and YEP medium for AGL1). The culture was then incubated on the incubator Shaker at 250 rpm at 30ºC for 3 hours. After that, the culture is spread on solid media that have been mixed with antibiotic selection (ampicillin 100 ppm for AGL1 and Streptomycin 50ppm for LBA4404).
2.3. Confirmation of Plasmids Carrying MeEF1A6 and CaMV35S Promoter in Agrobacterium tumefaciens Cultures
Plasmid confirmation was performed using the Polymerase Chain Reaction (PCR) method using specific primers to confirm that plasmid carrying MeF1A6 and CaMV35S promoter. The primers used are specific primers between the promoter base (forward) and the uidA gene coding base (GUS) (reverse). Specific primers were obtained from Apriyanto research [15]. The PCR condition follows used anealing temperature for MeEF1A6 promoters, which is 55 ºC and CaMV35S 60 ºC. MeEF1A6 bp fragment is 1311 bp, while CaMV35S is 675 bp. PCR results were obtained by running the electrophoresis method in agarose gel 1%. The DNA sequence tape was confirmed by visualization in UV light.
2.4. Introduction of MeEF1A6 promoter linked to the GUS gene into the tabacco plant
Introduction promoter was used by two methods, i.e., transient transformation and stable transformation. The introduction was carried out to investigate the work of the MeEF1A6 promoter in tobacco plants (N. tabaccum). A. Tumefaciens cultures that had been confirmed for the presence of pBI-MeEF1A6 and pBI-CaMV35S plasmids were sampled and transferred into liquid media containing the antibiotic selection. The culture was incubated at room temperature for 2-3 days. After incubation, 20 μL of liquid culture was transferred back to the liquid medium with a volume of 20 mL that had been added to selected antibiotics. The liquid culture is re-incubated for 12- 15 hours. After that, the culture was then confirmed OD600 until it reached 0.3-0.5. The culture was then centrifuged at 5000 rpm at 4ºC for 15 minutes. The supernatant was removed and the pellet was transferred to the infiltration media (MS0 with acetosyringone) which has been added with 20 ppm acetosyringone and 0.005% silwet.
The transient transformation plants prepared were separated per individual consisting of roots, stems, and leaves. The plants were then soaked in a co-cultivation solution with 20 ppm acetosiringone and siltettt 0.005% added. Then the infiltration stage was carried out using a vacuum and desiccator. The vacuum was done for 30 minutes. The infiltration process was carried out aseptically in the Laminar airflow cabinet (LAFC). Vacuumed plants in an infiltration medium then tightly closed in a container and incubated at 26ºC for 2-3 days. After 2-3 days, the plants are ready for further testing.
The stable transformation was carried out by following the method from Fajri (2015) [16]. The stable transformation begins with cutting 1x1 cm plant leaves. A total of 60 leaves that have been cut were immersed in the co-cultivation solutions containing A. tumefaciens and 20 ppm acetosyringone. The culture was then shaken with a 100 rpm speed shaker for 10 minutes. The culture was dried with sterile tissues for 10 minutes. After incubation, culture was then planted on solid infiltration media (Appendix A2). The abaxial position of the leaves is in the down position and submerged in solid co-cultivation media. The culture was then incubated in a dark room for 2-3 days. After incubation, the culture was washed with sterile deionized water 3 times and cefotaxime 200 ppm antibiotics 1 time. The culture was dried with sterile tissue and re-planted into callus growth media (Appendix A3). After two weeks, the culture was transferred to callus growth media that had been given antibiotic selection. Every 2 weeks the culture was transferred to a new callus growth media. The shoots that have emerged from the callus are transferred to the root induction medium (Appendix A4) containing selection antibiotics and left until the roots appear and become a new plantlet.
2.5. Analysis of uidA gene expression by Histochemical GUS Assay
Explants that had been incubated for 2 days and explants that had been successfully transformed were immersed in X-Gluc solution (1 mM 5-bromo-4-choloro-3-indolyl-β-Dglucuronide; Sigma), 100 mM sodium phosphate buffer pH 7 , 0.5 mM K3 [Fe(CN)6], 0.5 mM K4 [Fe(CN)6], 10 mM Na2EDTA, 0.1% (v / v), Triton X-100, then incubated at 37ºC for 18 hours in dark conditions. Then the explants were rinsed with 70% alcohol until the green color in the explants disappeared. Blue appearance was analyzed on the roots, stems, and leaves. The blue color indicated the promoter had been actived and stimulated expression of GUS gene that reacted with the substrate (X-Gluc) from colorless to blue. Samples that have been analyzed were then photographed using SLR (Canon EOS 1100 DC).
3. Results and discussion
3.1. Introduction and promoter sequence analysis
Introduction of promoters into A. tumefaciens was carried out by electroporation methods and selected by selection of antibiotics. The results of electroporation were performed by PCR test and obtained a band according to positive control. According to Apriyanto [15]. the CaMV35S :: uidA promoter PCR resulted in a base length of 675 bp fragment, while the MeEF1A6 :: uidA promoter produced a base length of 1311 bp fragment. The electrophoresis of AGL1 and LBA4404 PCR resulted containing CaMV35S promoters was showed band of about 675 bp fragment. These results are consistent with the positive control of the pBI121- CaMV35S plasmid. The electrophoresis of AGL1 and LBA4404 PCR resulted containing MeEF1A6 promoter was showed band about 1311 bp fragment. These results were also consistent with the positive control of the pBI121- MeEF1A6 plasmid (Figure 1). Therefore, it can be concluded that the plasmids pBI121-CaMV35S and pBI121-MeEF1A6 have been successfully introduced into A. tumefaciens strains AGL1 and LBA4404.
3.2. Qualitative Analysis of GUS activity with transien and stable transformation
The results of the introduction promoter with A. tumefaciens to tobacco plant (N. tabacum) was analyzed with histochemical GUS assay. The result from the histochemical GUS assay was showed a blue spot in the root, stems, and leaves in plants. The blue spot on the root was found in the transient transformation results using A. tumefaciens LBA4404 strain. The blue spot on leaves and stems was seen in the results of the transformation of the transient with both strains of A. tumefaciens (LBA4404 and AGL1) (Figure 2). Positive results were indicated by the blue spot in the explants. This was due to the association between MeEF1A6 and CaMV35S promoters with the uidA gene that expresses the β-glucuronidase enzyme. The uidA gene is located under the control of the MeEF1A6 promoter on the pBi121-MeEF1A6 plasmid and the CaMV35S promoter on the pBi121-CaMV35S plasmid. The expression of the uidA gene as a marker gene was controlled by the active promoters of MeEF1A6 and CaMV35S. The β-glucuronidase enzyme can hydrolyze the X-Gluc substrate which was previously a colorless compound to color (5.5'-dibromo-4,4' dichloro-indigo) blue with the help of K3Fe (CN)6 and K4Fe (CN)6 [17].
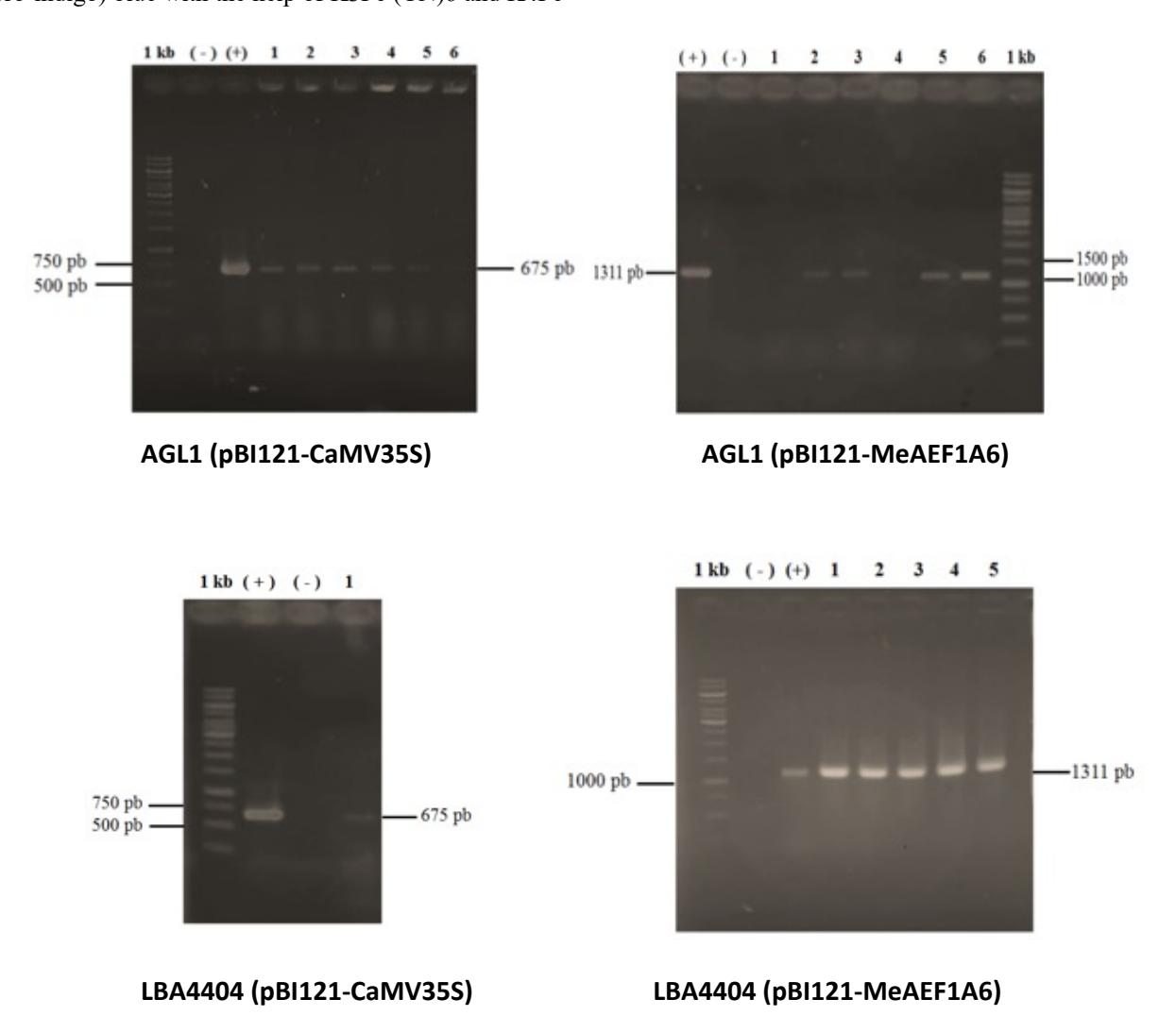
Figure 1. Confirmation PCR results of MeEF1A6 and CaMV35S promoter electroporation into Agrobacterium tumefaciens strain AGL1 and LBA4404. (+) shows positive control (plasmid containing promoter), (-) indicates negative control and number (1,2,3 etc) shows the number of samples tested
The transformation results using A. tumefaciens strain LBA4404 showed blue color on the roots, stems, and tobacco leaves. This is consistent with Apriyanto research [15] which shows that the MeEF1A6 promoter can work in all parts of tobacco sprouts. This proves that the MeEF1A6 promoter can activate the expression in roots, stems, and leaves. To see in more detail the specific work of the MeEF1A6 promoter, a cut on the trunk of the transient transformation results was made with LBA4404. Specimens of transient tobacco incision with MeEF1A6 promoter were compared with transient tobacco incision with CaMV35S promoter as positive control and wild-type tobacco incision as negative control (Figure 3).
The red circle shows a blue spot on the picture. The incision results show that the MeEF1A6 promoter can be expressed in the procambium tissue that lies between the xylem and phloem. This is different from the results of previous studies [18] which states that the color blue was found in the cortex, pith (not comprehensive), and xylem (a small part). In this study, blue also appeared in a small portion of the heartwood near procambium. The blue color in some of these tissues can prove that the MeEF1A6 promoter can work in tobacco plant tissue, although it is not comprehensive. The promoter MeEF1A6 and CaMV35S showed the same blue spot. This reinforces the notion that the MeEF1A6 promoter has the same constitutive nature as the CaMV35S promoter, which is a universal promoter in genetic engineering.
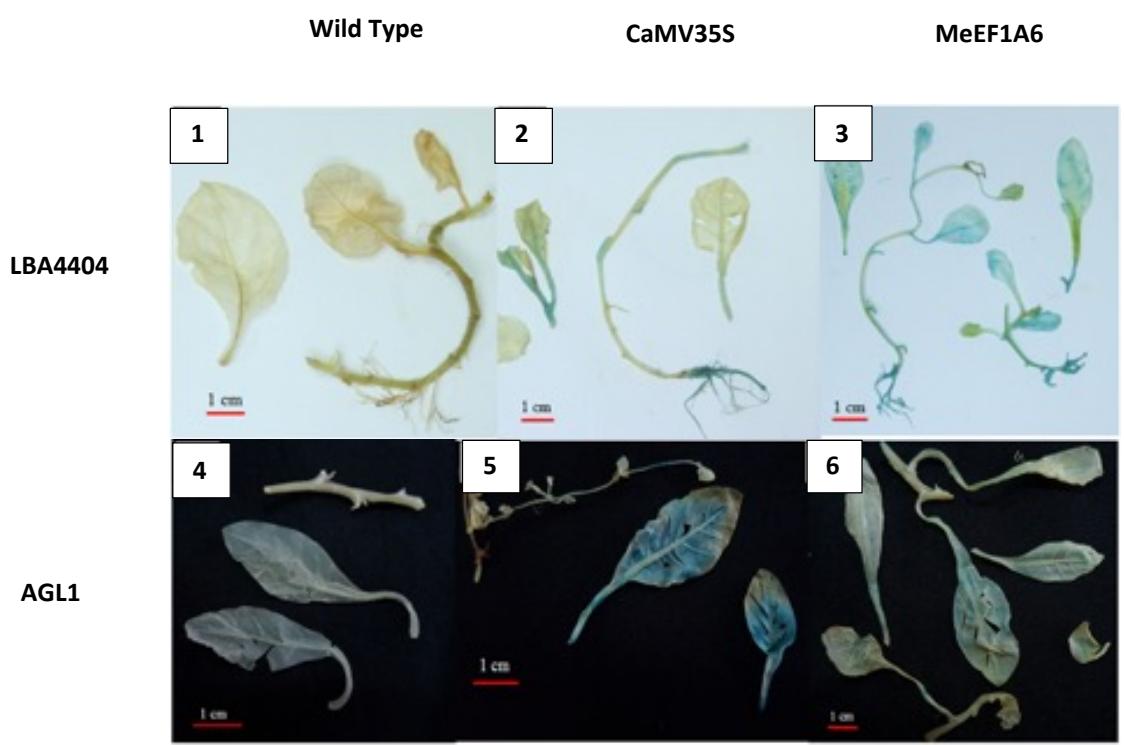
Figure 2. Transient transformation of tobacco plants (N.tabaccum) with A.tumefaciens strains AGL1 and LBA4404 which carry promoters MeEF1A6 and CaMV35S. (1-3) Showed transien transformation for LBA4404, (4-6) Showed transien transformation for AGL1.
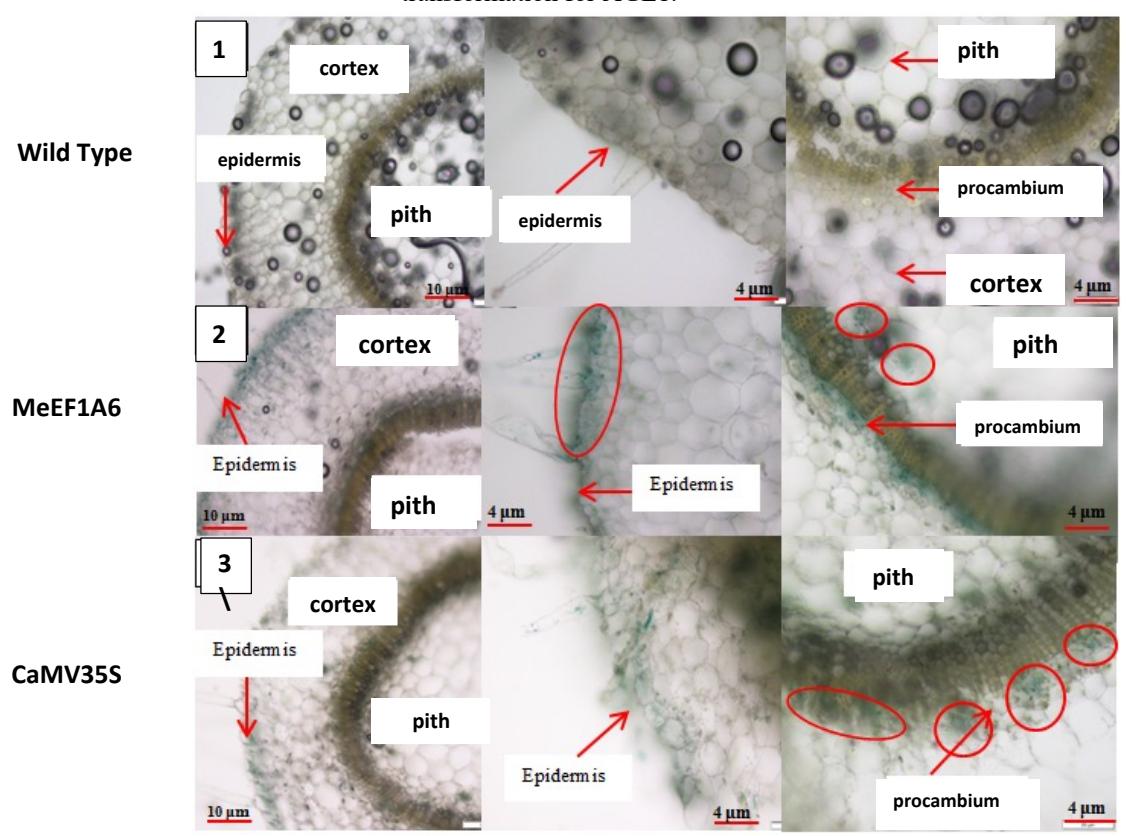
Figure 3. Cross-cutting incision of tobacco stem. (1) Wildtype tobacco, (2) LBA4404 transient transformation with pBI121-MeEF1A6,(3). LBA4404 transient transformation with pBI121-CaMV35S
Stable transformation using Agrobacterium strain AGL1 was obtained in the shoots, which then a GUS histochemical test was performed. In the stable transformation process, cefotaxime antibiotics are used to kill A. tumefaciens and kanamycin antibiotics to select explants. Cefotaxime 100 ppm antibiotics have been known to kill A. tumefaciens [16] and 50 ppm kanamycin antibiotics have been known to reduce plant growth [18]-[20]. The results of stable transformation with A. tumefaciens strain AGL1 can be seen in Table 1.
The stable transformation results with Agrobacterium strain AGL1 (Table 1) resulted in 12 living explants for pBI121- CaMV35S and 27 living explants pBI121-MeEF1A6. Live expansions have been inserted into the target promoter but it was uncertain whether they have been integrated into plant
DNA or not. Explants that had died were confirmed by looking at the yellowing of the explants that had not grown. The explants then performed a GUS histochemical test to see whether the promoters of MeEF1A6 and CaMV35S could work on stable transformation plants (Figure 4).
The blue color that appears indicates that the promoters CaMV35S and MeEF1A6 have successfully worked on tobacco plants that have been transformed. A study of Octacviana [18] showed that the blue color was very bright in almost all parts of the plant while in this study the blue color was only found in a few parts of the plant even though it included roots, stems, and leaves. To see more clearly whether the promoter is expressed in all types of plant tissue, a transverse incision is performed on the stem. The incision results can be seen in Figure 5.
| Table 1. The result of stable transformation of Agrobacterium strain AGL1 |
|---|
| Plants | Σ Leafs | Σ Leafs that had | Σ Shoots | Σ Explants | |
|---|---|---|---|---|---|
| been grown by callus | Dead | Live | |||
| pBI121-CaMV35S | 60 | 45 | 20 | 8 | 12 |
| pBI121-MeEF1A6 | 70 | 40 | 43 | 16 | 27 |
Figure 4. The result of stable transformation of Agrobacterium strain AGL1. (1) Tobacco wildtype, (2) pBI121- CaMV35S,(3) pBI121-MeEF1A6
Figure 5. Transverse incision resulting from stable transformation with Agrobacterium strain AGL1; a. pBI121-CaMV35S; b. pBI121-MeEF1A6
Figure 5 showing that the MeEF1A6 promoter can work in the procambium, xylem, and some pith tissue. This is consistent with the results of research before [18] who also stated that the MeEF1A6 promoter can work in the cortex, xylem, and pith network. In this study, the blue color did not spread evenly to the cortex. This also happened to the positive control (CaMV35S). This is possible because the results of the stable transformation are still not evenly distributed in all host cells or chimeras / chimeric. Chimera is a condition where individuals have different genomes in several cells, resulting in variations in expression/phenotypes in one individual [21]. So the MeEF1A6 promoter is only expressed in a few cells/tissues and cannot be maximally expressed in GMO tobacco plants.
4. Conclusion
Promoter MeEF1A6 can actively work on tobacco plants (N. tabacum), though not showing high activity. This study proved that the promoter MeEF1A6 could work in several tobacco plant tissues; procambium, epidermis, cortex, and pith. This adds to the suspicion that the MeEF1A6 promoter can work constitutively on tobacco plants of SR1 variety (Nicotiana tabacum cv SR1).
Acknowledgements
The authors thanked the Genetic research group of SITH ITB and PPSHB IPB for all supports and facilities that support this research. The authors also thanked Ian for helping during the research in the laboratory.