
1. Introduction
Energy is a pivotal necessity impacting various aspects of daily life, including fuel for machinery and transportation. Currently, 85% of global energy consumption relies on nonrenewable resources, making the fuel industry vulnerable to energy crises [1]. Consequently, alternative energy from renewable resources is urgently needed. Biodiesel emerges as a promising renewable energy source, integrating a neutral carbon cycle and potentially addressing issues related to conventional diesel consumption.
Biodiesel can be produced using oils or lipids from organisms such as Chlorella vulgaris. Microalgae offer several advantages over conventional terrestrial biomass like corn, as they can be cultivated year-round, leading to higher biomass productivity, short growth cycles, high lipid yields, efficient CO2 fixation, and minimal land usage without competing with food resources [2].
One major challenge in lipid extraction is the low yield
due to the complex and rigid cell wall structure of microalgae, which hinders solvent interaction with intracellular lipids and necessitates large volumes of solvent [3]. Cell wall disruption is essential to maximize lipid yield. However, conventional disruption methods, such as chemical treatments with ionic liquids or physical methods like sonication, have drawbacks, including high energy consumption, high costs, complex processes, and environmental concerns [4, 5].
A promising alternative is the use of deep eutectic solvents (DES), environmentally friendly green solvents, as a pretreatment to overcome the limitations of conventional cell disruption and solvent extraction methods. DES are simple to prepare, cost-effective, non-toxic, and biodegradable [6]. They are typically synthesized from organic salts like choline chloride or choline acetate and hydrogen bond donors such as carboxylic acids, amides, or amines to form eutectic mixtures with lower melting points than their individual components [7]. Natural deep eutectic solvents (NaDES) are a subset of
DES synthesized from compounds found in plant primary metabolites, such as organic acids, sugars, and choline derivatives [8].
NaDES can influence the hydrogen bonds within the microalgae cell wall, thereby affecting its integrity and permeability [3]. Factors influencing the efficiency of NaDES in extraction or cell disruption include the ratio of organic salt to hydrogen bond donor, the microalgae strain, and the amount of deionized water added for aqueous DES. This study aims to analyze the impact of NaDES application on the microalgae Chlorella vulgaris to achieve efficient lipid production with high yields for biodiesel precursors.
2. Methodology
2.1. Microalgae culture
The microalgae culture used in this research was Chlorella vulgaris, obtained from The Laboratory of Indonesian Culture Collection (InaCC) in Cibinong. The cultivation medium was commercial NPK fertilizer (Nutricomp-D), which contains 22% total nitrogen (N), 17% phosphorus pentoxide (P₂O₅), 15% potassium oxide (K₂O), and 12.5% magnesium oxide (MgO). Other chemicalsfor NaDES synthesis, such as choline chloride (ChCl), glycerol, and oxalic acid, were sourced from Merck and HiMedia.
2.2 Synthesis of Natural Deep Eutectic Solvents (NaDES)
NaDES were synthesized using choline chloride (ChCl) as the hydrogen bond acceptor (HBA) and glycerol or oxalic acid as the hydrogen bond donors (HBD). The molar ratios of the mixtures were varied as 1:1 and 1:2 (HBA:HBD) for each NaDES variation. ChCl, glycerol, and oxalic acid were weighed according to the respective molar ratios and added to an Erlenmeyer flask. The mixtures were stirred at 80°C using a magnetic stirrer until a colorless solution was obtained (after 1-2 hours). The NaDES solutions were then stored in an oven at 50°C for 24 hours to prevent water absorption and subsequently kept in tightly closed falcon tubes in a desiccator before use. Before use in the extraction process, NaDES (ChCl-G and ChCl-OA) were mixed with 40% (v/v) deionized water to decrease viscosity and enhance mass transfer.
2.3 Biomass Pre-treatment with NaDES
Dried Chlorella vulgaris biomass was pre-treated with NaDES molar variations: ChCl-G (1:1), ChCl-G (1:2), ChCl-OA (1:1), and ChCl-OA (1:2). The dried microalgae were mixed with NaDES at a solvent-to-biomass ratio of 5 mL/g. Pre-treatment was conducted at room temperature (25-27°C) for 24 hours with constant stirring using a magnetic stirrer. Post-treatment, NaDES and biomass were separated via centrifugation at 6,000 rpm for 20 minutes. The supernatant containing some lipids mixed with NaDES was stored in a desiccator before GC-MS analysis. The pre-treated biomass was washed with deionized water and centrifuged four times at 6,000 rpm for 10 minutes each. The biomass pellet was dried in an oven at 60°C for 24 hours, then ground into a powder using a mortar and pestle for further analysis.
2.4 Lipid Extraction using Bligh-Dyer and Hexane Methods
Lipid extraction from dried microalgae biomass was performed using the Bligh & Dyer method (1959) as a control to compare traditional solvent extraction (chloroformmethanol) with the NaDES pre-treatment followed by hexane extraction [9]. This control method helps evaluate the effectiveness of NaDES in improving lipid yield relative to a widely used conventional extraction technique. Dried biomass ( 1 gram) was mixed with 60 mL of chloroform and methanol (1:2 v/v) in a Soxhlet system for 5 hours at 70°C. Following this, 1 mL of chloroform and 1 mL of water were added to the solution and vortexed for 1 minute. The mixture was centrifuged at 5,000 rpm for 8 minutes, separating the organic phase containing lipids and chloroform. Dried pretreated microalgae with NaDES were also extracted using the same method but with hexane as the solvent. NaDES pre-treated microalgae were extracted using hexane, chosen for its relevance to industrial lipid extraction. The Bligh-Dyer method (chloroform-methanol) was used as a control to compare traditional and industrially relevant solvents. Both methods provide comparable fatty acid profiles for GC-MS analysis, validating the comparison. The mixture of microalgae lipid and solvent was evaporated on a water bath at 60°C for approximately 2 hours to obtain the lipid content. Lipid yield was determined using the following equation:
\[Y_{lipid} = \frac{W_{lipid}}{W_{dry\ biomass}} \times 100\%\] (1)
in which Ylipid is lipid yield (% w/w), Wlipid is extracted lipid weight (gr), and Wdry is the weight of dried biomass (gr).
2.5 Determination of Acid Value and Free Fatty Acid Content
The acid value and free fatty acid (FFA) content were determined using the titration alkalimetry method by a nationally accredited reference laboratory. Lipid samples (0.3 g) were weighed into a 250 mL Erlenmeyer flask and diluted in ethanol solvent, with 3 drops of phenolphthalein (PP) indicator added. The prepared sample was titrated against 0.02 N KOH solution until a pink color persisted for 30 seconds. The acid value was calculated using the following equation:
\[AV = \frac{V_{titration} x M r_{KOH} x N_{KOH}}{W_{sample}}\] (2)
In which AV is acid value, Vtitration is titration volume (ml), MrKOH is the molecular weight of KOH which is 56,11 gr/mol, NKOH is KOH concentration which is 0,02 N, and Wsample is lipid weight (gr).
Free fatty acid (FFA) content was considered as palmitic acid. Lipid samples (0.5 g) were weighed and diluted with ethanol in a 250 mL Erlenmeyer flask, with 3 drops of PP indicator added. The prepared sample was titrated against 0.02 N NaOH solution until a pink color persisted for 30 seconds. The FFA content was calculated using the following equation:
\[FFA = \frac{V_{titrasi} \times Mr_{PA} \times N_{NaOH}}{W_{sampel} \times 1000} \times 100\% \quad (3)\]
In which FFA is free fatty acid content (%), Vtitrasi is titration volume (mL), MrPA is palmitic acid molecular weight which is 256 gr/mol, NNaOH is NaOH concentration which is 0,05 N, dan Wsample is lipid weight (gr).
2.6 Fatty Acid Components Analysis
The fatty acid profile of Chlorella vulgaris lipid was analyzed using gas chromatography-mass spectrometry (GC-MS) with a flame ionization detector (FID) by a nationally accredited reference laboratory. The lipid extract used for GC-MS analysis was obtained from the pooled extract of both the colloidal phase and the biomass extract. Lipid samples were filtered using a 0.22 µm syringe filter. The stationary phase was cyanopropyl on a DB FastFame capillary column. The injector temperature was set at 300°C, and the detector temperature was 250°C. Argon gas was used as the mobile phase with a flow rate of 1 mL/min. The oven column's initial temperature was 190°C for 5 minutes, followed by a heating rate of 10°C/min until 250°C. The temperature was maintained at 250°C for 5 minutes, then increased to 280°C at a heating rate of 10°C/min, held for 30 minutes, and finally cooled to room temperature.
2.7 Statistical Analysis
Experiments were conducted in duplicate, and mean values were reported. Data processing was performed using Microsoft Excel (Office Enterprise 2019) with single-factor analysis of variance (ANOVA) to determine statistical significance.
2.8 Manuscript Preparation
In preparing this manuscript, the authors utilized several AI-based tools to enhance the quality and clarity of the writing. ChatGPT4.0 was employed to improve language clarity and ensure concise, precise communication of scientific concepts. For efficient and accurate reference management, Zotero was used to organize and cite all sources. Grammarly assisted in refining the English language and ensuring adherence to proper grammar and style, which is crucial for clear communication with an international audience. Additionally, QuillBot was applied for paraphrasing certain sections, helping to maintain originality while preserving the intended meaning of rephrased content.
3. Results and Discussion
3.1. Chlorella vulgaris Microalgae Growth Curve and Biomass Productivity Analysis
The growth curve of Chlorella vulgaris biomass is presented in Figure 1, showing multiple growth phases: lag, log (exponential), stationary, and death phases. The lag phase occurred after biomass inoculation until the third day of cultivation, during which microalgae acclimatized to new conditions such as medium pH, temperature, and light [10].
A slight decrease in biomass was observed on the first day after inoculation, with cell density dropping by 1% compared to the immediate post-inoculation value. Generally, biomass decline for Chlorella vulgaris during the lag phase ranges from 3.81% to 4.5% [10]. In this study, the cell decline was lower than literature values, possibly due to the use of
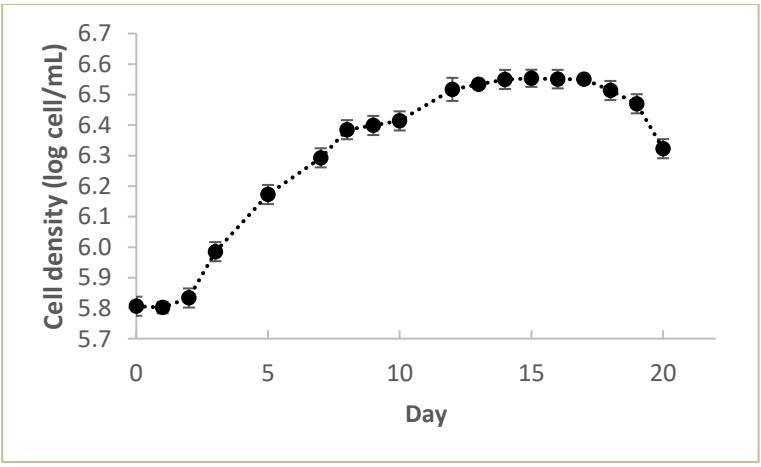
Figure 1. Chlorella vulgaris biomass growth curve
similar medium and cultivation conditions, facilitating faster acclimatization.
The log phase, characterized by active cell division and biomass increment, spanned from day 3 to day 13. The specific growth rate was determined based on biomass at the beginning and end of the log phase. The stationary phase, where growth and death rates equalize, occurred from day 13 to day 17, with maximum biomass concentration of 0.057 g/L and cell density of 3.54 x 10⁶ cells/mL reached on day 14. The death phase from day 18 to day 20 was marked by a drastic decline in cell density due to reduced nutrient availability, dissolved oxygen, and potential contamination [11].
The specific growth rate (µ) of Chlorella vulgaris in this study was 0.127 day⁻¹, with a doubling time of 5.433 days. Literature reports a specific growth rate of 0.344 day⁻¹ after 7 days of cultivation with a maximum cell density of 1.88 x 10⁷ cells/mL using pure CO₂ aeration [12]. Variations in growth rates are attributed to differences in microalgae strain, medium-induced stress, and cultivation conditions.
Pure CO₂ aeration, for instance, increases CO₂ fixation rates, thereby enhancing metabolic reactions and doubling the growth rate compared to regular aeration (0.04% CO₂) [13]. This study's doubling time of 5.433 days for Chlorella vulgaris with regular aeration contrasts with the 2.015 days reported for pure CO₂ aeration [12].
Biomass productivity in this study was 0.004 g·L⁻¹·day⁻¹, lower than the general biomass productivity range of 0.025 - 0.040 g·L⁻¹·day⁻¹ reported in other studies [14, 15]. Differences in cultivation treatments, such as medium composition, contribute to this variation. In literature, media like F2 and Nutriverde with higher nitrogen concentrations (9.29% and 5.571%, respectively) were used, while the NPK Nutricomp-D commercial fertilizer used in this study contained only 2.2% nitrogen. Thus, lower nitrogen availability in this study led to lower maximum biomass and productivity.
3.2. Lipid Character Analysis of Chlorella vulgaris Based on Acid Value and Free Fatty Acid Content
Analyzing the lipid characteristics of Chlorella vulgaris is crucial for determining its suitability for biodiesel production. This study evaluated lipid character based on acid value and free fatty acid (FFA) content. The acid value indicates the amount of potassium hydroxide (KOH) required to neutralize free fatty acids in the lipid. The acid value in this study was 42.56 mg KOH/g lipid.
Animal fats and vegetable oils used as biodiesel precursors typically have acid values between 50 - 200 mg KOH/g [16, 17, 18]. Chlorella vulgaris lipids generally have acid values ranging from 25 - 130 mg KOH/g [17, 18], aligning with the values observed in this study. However, the acid value remains high compared to biodiesel standards, indicating potential corrosiveness towards materials like aluminum, steel, and nickel [18]. High acid values can also lead to saponification
during transesterification, reducing biodiesel yield and complicating product separation [18].
International standards such as ASTM D6751 and EN14214 require acid values below 0.5 - 0.8 mg KOH/g for biodiesel. Post-transesterification, biodiesel typically exhibits lower acid values, such as 0.6 mg KOH/g, indicating the potential for reduced acid values following transesterification and purification processes [19].
FFA content, indicative of the lipid's energy reserve in microalgae cells and formed through triglyceride hydrolysis, influences the lipid's oxidative stability and potential for biodiesel conversion. High FFA content suggests susceptibility to oxidation and potential clogging of machinery fuel filters, especially at lower temperatures [20]. Biodiesel production generally requires lipids with FFA content below 0.5% (w/w) [21].
In this study, the FFA content of Chlorella vulgaris lipid was 0.25% (w/w), indicating compatibility with biodiesel production conditions. The cultivation treatment used in this study resulted in lower FFA content compared to other studies, which reported FFA contents ranging from 0.5 - 6.9% (w/w) [21, 22, 23]. The lower FFA content observed may be attributed to the use of NPK Nutricomp-D, a minimal medium with reduced nitrogen concentration (0.158 g/L) compared to Bold Basal Medium (BBM) used in other studies (0.25 g/L). This reduced nitrogen concentration likely shifted the lipid composition towards higher triglyceride content and lower FFA levels, which is more suitable for biodiesel production [24].
3.3. Analysis of NaDES Application Influence on Microalgae Chlorella vulgaris Lipid Yield
The efficiency of lipid extraction from microalgae can be significantly affected by the solvent and pre-treatment methods used. In this study, the application of natural deep eutectic solvent (NaDES) as a pre-treatment method showed a positive impact on lipid yield from Chlorella vulgaris. Multiple variations of NaDES, consisting of choline chloride (ChCl) combined with glycerol (ChCl-G) or oxalic acid (ChCl-OA), and varying molar ratios, were tested.
The total lipid yield included lipids from both the colloid phase and the dry biomass post-NaDES pre-treatment. When the biomass was agitated in NaDES solution for 24 hours, lipids were released from the microalgae cells into the NaDES phase due to the cell disruption process. Additionally, since NaDES also functions as a liquid solvent, it facilitated lipid extraction during the disruption process. The lipid content extracted during the disruption was dispersed and mixed with the NaDES, forming the colloid phase.
Subsequent drying and extraction of the biomass post-pretreatment using hexane yielded further lipids, contributing to the total lipid yield. Figure 2 illustrates the total lipid yields for each NaDES variation.
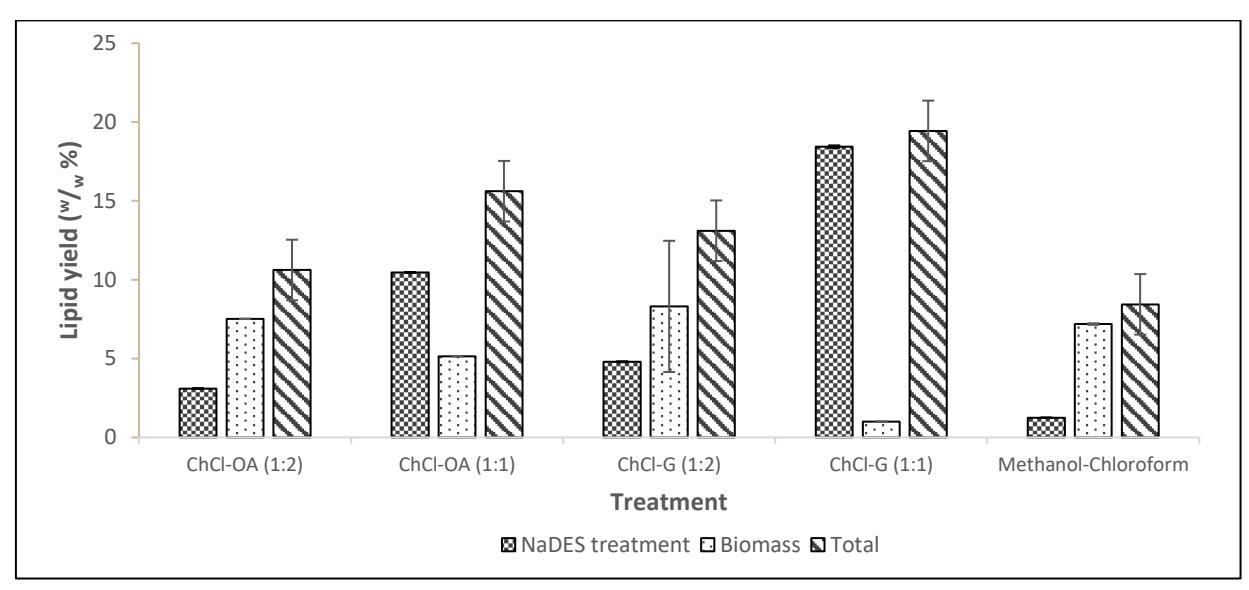
Figure 2. Lipid yield from the colloidal phase of NADES pre-treatment and dried biomass
The application of NaDES as a pre-treatment method significantly improved total lipid yields compared to untreated biomass, which had a total lipid yield of 8.44% (w/w). The highest total lipid yield, achieved using the NaDES mixture of ChCl and glycerol with a 1:1 molar ratio, reached 19.44% (w/w), more than doubling the yield from untreated biomass. In contrast, the lowest lipid yield was observed with the NaDES mixture of ChCl and glycerol at a 1:2 molar ratio. Overall, every NaDES and molar ratio variation experienced an increase in lipid yield compared to the untreated sample, demonstrating the effectiveness of NaDES pre-treatment in enhancing lipid extraction efficiency.
Literature indicates that NaDES pre-treatment with ChCl-OA (1:2), as well as urea and acetamide (1:2), could increase lipid yields by up to 80.90% and 75.25%, respectively, compared to untreated biomass [3]. The statistically significant p-values (p < 0.05) for all NaDES and molar ratio variations support the hypothesis that NaDES application can enhance lipid yield and extraction efficiency in microalgae Chlorella vulgaris.
NaDES solution is synthesized by combining the hydrogen bond acceptor (HBA) and hydrogen bond donor (HBD). This reaction not only forms hydrogen bonds within the NaDES but also allows these components to interact with the microalgae cell wall, facilitating hydrogen bonding there as well [25]. This interaction enhances the rigidity and integrity of the cell wall, thus improving the release of intracellular lipids due to increased cell wall permeability—a result of the energy reduction in hydrogen bond interactions with macromolecules and hemicellulose [3, 7]. Consequently, NaDES application significantly boosts the efficiency of the lipid extraction process by improving solvent access to intracellular lipids [26].
The hydroxyl and carboxyl groups in NaDES form bonds with similar groups in the cell wall, impacting the hydrogen
bonds between cellulose and hemicellulose that typically confer rigidity to microalgae cells [25]. Continuous stirring for 24 hours during the treatment also assists in the cell disruption process, enhancing the solvent's access to the cell wall and facilitating interaction with intracellular lipids.
According to this study, NaDES formulations comprising choline chloride (ChCl) and glycerol yielded higher total lipid outputs compared to those containing ChCl and oxalic acid. This variation in lipid yield may be attributed to differences in solvent viscosity. Generally, NaDES is characterized by its high viscosity, particularly evident in mixtures of ChCl and oxalic acid, which are composed of two solid compounds. The viscosity of the NaDES ChCl-OA (1:2) is 212.9 mm²/s, which is significantly higher compared to NaDES ChCl-G (1:1) at 52.3 mm²/s [27]. The elevated viscosity in the ChCl-OA mixture could restrict the solubility of microalgae during the extraction process, making it more challenging for the solvent to interact with the microalgae cell surface due to increased surface tension [28].
Comparative analysis of NaDES formulations shows that those based on ChCl and glycerol yield higher total lipid outputs compared to those with ChCl and oxalic acid, likely due to differences in solvent viscosity. The addition of 40% deionized water was employed to lower the viscosity of NADES, as high viscosity can hinder mass transfer during extraction. By reducing the viscosity, the NADES mixture became less resistant to flow, which allowed for improved solvent penetration into the microalgae cell wall. This enhanced solvent access to intracellular lipids, thereby increasing the overall extraction efficiency. However, care was taken to ensure that the addition of water did not dilute the NADES excessively, as this could disrupt the eutectic structure and reduce its efficacy as a cell wall disruptor. Adjusting the NaDES viscosity by adding deionized water reduces this effect, although excessive water can disrupt
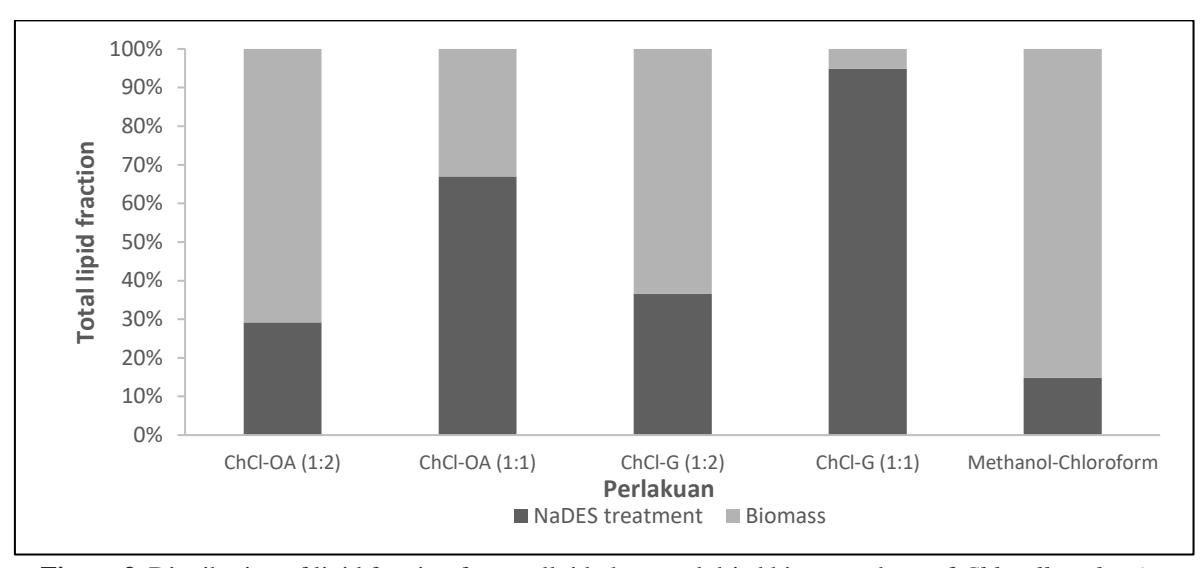
Figure 3. Distribution of lipid fraction from colloid phase and dried biomass phase of Chlorella vulgaris
the hydrogen bonding in NaDES, potentially degrading its effectiveness [28].
The molar ratio of HBA to HBD significantly influences the lipid yield, with a 1:1 ratio proving most efficient. This efficiency is reflected in the lipid yields from both the colloid and dried biomass phases. Notably, the highest lipid yield from the colloid phase was observed with NaDES ChCl-G (1:1), indicating superior permeability enhancement of the microalgae cell wall [27]. In contrast, the lipid yield from dried biomass after NaDES treatment shows variability, suggesting that further extraction from the biomass is necessary even after initial NaDES treatment.
The lipid yields from the colloid phase and dried biomass exhibit distinct variations across different NaDES treatments. The highest lipid yield from the colloid phase was observed in the NaDES formulation of ChCl-G (1:1), achieving 18.44% (w/w). This high yield underscores the effectiveness of ChCl-G in enhancing the permeability of microalgae cell walls, thereby facilitating the release of lipid content into the solvent during treatment. In contrast, the lowest lipid yield in the colloid phase, at only 1.25% (w/w), was found in the untreated samples.
The lipid yield from dried biomass after pre-treatment also concur some differences for each variation. The highest lipid yield is found in ChCl-G (1:2), which is 8,31% (w/w). Meanwhile, the lowest lipid yield is found on ChCl-G (1:1), which is 1% (w/w). The lipid yield from biomass shows that after NaDES pre-treatment, that the dried biomass potentially still has lipid content even though the cell wall has experienced lysis. Therefore, further biomass extraction after NaDES pretreatment is still needed.
There are observable trends in the distribution of lipid yields between the colloid phase and dried biomass. Samples treated with NaDES generally exhibit higher lipid yields from the colloid phase, as demonstrated in Figure 3. This trend suggests that during NaDES application, intracellular
lipids are effectively mobilized from the cell wall matrix into the solvent under continuous mixing conditions. Although methanol-chloroform was used as a control without NADES pre-treatment, a small colloidal phase was observed. This is likely due to the presence of some emulsified lipids in the solvent, which formed a colloidal suspension during the extraction process. The phenomenon has been noted in other studies where organic solvent extraction can result in the formation of colloidal phases even without prior cell disruption. The methanol-chloroform extraction method yielded the lowest lipid recovery, likely due to its inability to disrupt the rigid cell wall of Chlorella vulgaris as effectively as the NADES pre-treatment. While methanol-chloroform is a standard solvent for lipid extraction, it does not promote cell wall permeability, resulting in lower lipid release compared to the enhanced extraction achieved with NADES, which facilitates cell disruption
Moreover, it is evident that when a majority of lipids are released during NaDES pre-treatment, the residual lipid content in the dried biomass is reduced. This observation is supported by a strong negative correlation coefficient of -0.90 between lipid yields from the biomass and those from the colloid phase [3, 27]. This correlation indicates that higher lipid yields in the colloid phase correspond to lower yields in the dried biomass. This relationship is also noted in other studies [3], which report that the decline in lipid content from dried pre-treated biomass is attributable to the migration of lipids from the cell membrane into the NaDES aqueous phase.
3.4. NaDES Application Effect Towards Fatty Acid Profile of Microalgae Chlorella vulgaris Biomass
The quality of biodiesel derived from the transesterification of microalgae lipids is largely influenced by the characteristics of the triglycerides, which depend on the distribution and composition of the fatty acids in the lipid. The transesterification reaction, which converts lipids into fatty
Fatty Acid Fatty Acid Components (% w/w) ChCl-OA (1:2) ChCl-OA (1:1) ChCl-G (1:2) ChCl-G (1:1) Untreated Biomass ChCl-G (1:2) C12:0 - 0,69 ± 0,005 - 0,77 ± 0,014 - - C14:0 0,07 ± 0,00 0,38 ± 0,005 - 0,53 ± 0,013 - - C16:0 0,92 ± 0,022 3,18 ± 0,055 1,05 ± 0,00 4,35 ± 0,061 0,286 ± 0,004 2,42 ± 0,030 C16:1 - - - - - 1,02 ± 0,013 C18:0 0,27 ± 0,005 0,74 ± 0,013 0,35 ± 0,008 1,13 ± 0,016 0,17 ± 0,001 0,20 ± 0,005 C18:1 1,29 ± 0,028 4,38 ± 0,078 2,11 ± 0,049 7,79 ± 0,144 0,49 ± 0,012 1,49 ± 0,035 C18:2 0,45 ± 0,011 1,11± 0,019 1,10 ± 0,023 2,20 ± 0,044 0,15 ± 0,00 2,86 ± 0,055 C18:3 - - 0,19 ± 0,004 1,64 ± 0,028 0,16 ± 0,002 2,51 ± 0,042 C20:2 - - - - - 0,75 ± 0,016 MUFA 1,29 ± 0,028 4,38 ± 0,078 2,11 ± 0,049 7,79 ± 0,1442 0,49 ± 0,012 2,51 ± 0,0221 PUFA 0,60 ± 0,014 1,11 ± 0,019 1,30 ± 0,027 3,84 ± 0,028 0,31 ± 0,002 6,15 ± 0,0419
TOTAL 3,15 ± 0,07 10,47 ± 0,18 4,80 ± 0,090 18,40 ± 0,320 1,25 ± 0,020 11,25 ± 0,17
Table 1. Fatty acid components of lipid from colloid phase and dried biomass
acid methyl esters (FAME) or biodiesel, does not alter the fatty acid composition of the lipid precursors. Consequently, the fatty acid components play a critical role in determining the biodiesel's quality, affecting properties such as cetane number and cold flow [29].
Fatty acid composition is analyzed using gas chromatography-mass spectrometry (GC-MS), with results for each variation displayed in Table 1. The fatty acid content, expressed as a percentage of the dry biomass weight, is sampled from the colloid phase after NaDES pre-treatment. Additionally, pre-treated dried biomass samples, particularly for the ChCl-G (1:2) variation, are analyzed for fatty acid content.
Detected fatty acid types range from C12 to C20. Predominantly, the fatty acid composition is dominated by C16 (palmitic acid) and C18 (stearic, oleic, linoleic, and linolenic acids). This dominance is consistent with other studies, which frequently report high concentrations of C16 and C18 fatty acids in Chlorella vulgaris [3]. The high content of these fatty acids is attributed to their solubility in the solvents used, which facilitates greater extraction efficiency compared to other fatty acids.
Table 2 illustrates the fraction distribution for each fatty acid relative to the total lipid content, providing a comparison with literature references for untreated biomass. This data reveals that C16 and C18 fatty acids contribute to 80-90% of the total fatty acid content, significantly influencing the biodiesel's final characteristics and quality. The highest fraction distribution of C16 is observed in the colloid phase of the NaDES ChCl-OA (1:1) variation, at 30.35%, while the highest distribution of C18 is found in the colloid phase of ChCl-G (1:2), at 74.13%.
The prominent presence of C18 and C16 fatty acids is advantageous as they enhance fuel quality and ignition properties. Additionally, a high concentration of C18 fatty acids contributes to greater oxidative stability during storage. Saturated fatty acids, such as palmitic and stearic acid, are associated with higher cetane numbers, suggesting that the NaDES ChCl-OA (1:1) colloid phase sample exhibits significant potential as a biodiesel precursor, meeting the EN14214 standard's minimum cetane number requirement of 51. In contrast, untreated biomass samples contain longer fatty acid chains like C22:6 and C20:5, which are absent in the studied samples and are known to adversely affect biodiesel quality by lowering the cetane number and promoting fuel oxidation [30].
Figure 4 provides a comparison of the fraction distribution of polyunsaturated fatty acids (PUFA), monounsaturated fatty acids (MUFA), and saturated fatty acids (SFA) for each variation. The ChCl-G (1:2) biomass sample exhibits a higher PUFA distribution, leading to a lower cetane number and higher iodine number, indicating potential issues with smoking upon ignition and reduced oxidation stability. Conversely, the ChCl-OA (1:1) colloid phase sample, with a lower PUFA distribution, is likely to exhibit higher oxidation stability and a lower iodine number, aligning more closely with international biodiesel standards, which limit C18:3 distribution to a maximum of 12% to avoid undesirable properties in biodiesel precursors.
The lipid composition rich in long-chain saturated fatty acids typically leads to a lower cold filter plugging point, causing precipitation and potential filter clogging. However, this study did not detect long saturated fatty acid chains. It is observed that the colloid phase samples ChCl-G (1:2) and ChCl-OA (1:1) have a higher distribution of MUFA, particularly C18:1 (oleic acid), suggesting that these lipids
Table 2. Fraction distribution for each fatty acid contents
| Fatty Acid | Fraction Distribution for Each Fatty Acids (%) | ||||||
|---|---|---|---|---|---|---|---|
| ChCl-OA (1:2) | ChCl-OA (1:1) | ChCl-G (1:2) | ChCl-G (1:1) | Untreated | Biomass ChCl-G (1:2) | Biomass untreated [35, 36] | |
| C4:0 | - | - | - | - | - | - | 0,19 % |
| C6:0 | - | - | - | - | - | - | 0,14% |
| C8:0 | - | - | - | - | - | - | 0,13% |
| C10:0 | - | - | - | - | - | - | 0,19% |
| C12:0 | - | 6,58% | - | 4,18% | - | - | |
| C14:0 | 2,21% | 3,61% | - | 2,85% | - | - | 1,3% |
| C15:0 | - | - | - | - | - | - | 0,19% |
| C16:0 | 29,19% | 30,35% | 21,84% | 23,62% | 22,88% | 21% | 29,52% |
| C16:1 | - | - | - | - | - | 9% | 1,88% |
| C17:0 | - | - | - | - | - | - | 0,24% |
| C17:1 | - | - | - | - | - | - | 6,00% |
| C18:0 | 8,53% | 7,04% | 7,24% | 6,12% | 13,38% | 2% | 2,74% |
| C18:1 | 40,90% | 41,80% | 43,88% | 42,35% | 39,22% | 13% | 17,01% |
| C18:2 | 14,41% | 10,63% | 23,01% | 11,95% | 11,83% | 25% | 27,74% |
| C18:3 | - | - | 4,03% | 8,92% | 12,69% | 22% | 11,5% |
| C20:2 | - | - | - | - | - | 7% | - |
| C20:4 | 5% | - | - | - | - | - | - |
| C20:5 | - | - | - | - | - | - | 0,13% |
| C22:6 | - | - | - | - | - | - | 0,13% |
| SFA | 39,93% | 47,57% | 29,03% | 36,98% | 36,26% | 23% | 64,39 |
| UFA | 60,07% | 52,43% | 70,92% | 63,22% | 63,74% | 77% | 35,61 |
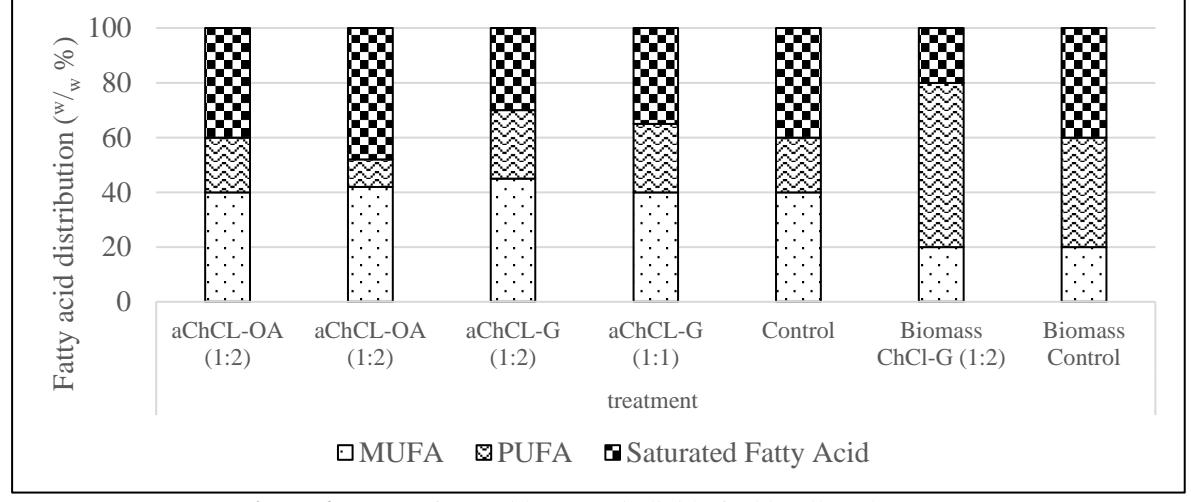
Figure 4. Omega fatty acid content in lipid of Chlorella vulgaris
may possess favorable general biodiesel characteristics and are likely to meet global biodiesel standards. Therefore, NaDES pre-treatment could potentially aid in ensuring compliance with standard biodiesel properties, enhancing the viability of microalgae-derived biodiesel.
According to the GC-MS results, the extracted lipid from Chlorella vulgaris includes significant quantities of omega fatty acids, as detailed in Figure 5. The omega fatty acid content within these lipids shows promising potential for use as pharmaceutical feedstocks, such as health supplements. This is particularly relevant for applications where the fatty acid characteristics are less suitable for biodiesel production. The omega fatty acids present, specifically omega-3, omega-6, and omega-9, are essential fatty acids that cannot be synthesized by the human body and are predominantly derived from polyunsaturated fatty acids (PUFAs), making them highly valuable for supplement production in the pharmaceutical industry [31].
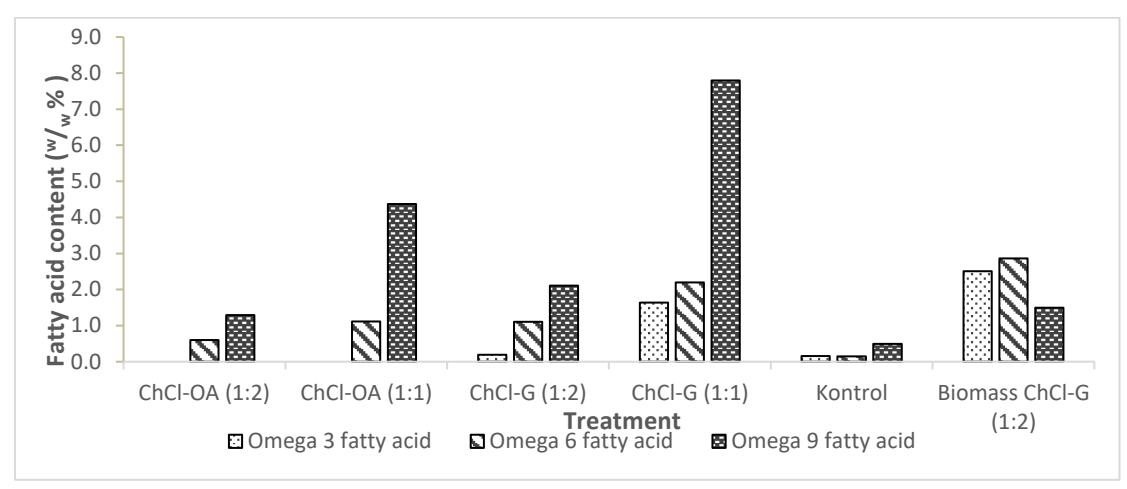
Figure 5. Omega fatty acid content in lipid of Chlorella vulgaris
In particular, the lipid samples from the dried biomass variation ChCl-G (1:2) and the colloid phase ChCl-G (1:1) exhibit the highest contents of omega fatty acids. The omega-6 content in these variations is measured at 2.86% and 2.199% respectively, while omega-3 content is found to be 2.51% and 1.6416% respectively. These concentrations indicate a robust potential for these specific lipid extracts to be developed into nutritional supplements, providing an alternative application for microalgae lipids that extends beyond energy production into health and wellness sectors.
The colloid phase sample of ChCl-OA (1:2) exhibits a lower overall omega fatty acid content but notably includes C20:4 omega-6, a long-chain fatty acid. Long-chain fatty acids, while beneficial in various applications, are not ideal as biodiesel precursors due to their impact on fuel properties such as viscosity and cold flow behavior. Consequently, this specific lipid composition makes the ChCl-OA (1:2) sample more suitable for pharmaceutical applications, particularly in the production of omega supplements, where long-chain omega-6 fatty acids are highly valued for their health benefits.
Conversely, the colloid phase sample of ChCl-G (1:1) contains higher contents of omega-3 and omega-6 fatty acids, which, combined with a higher monounsaturated fatty acid (MUFA) and omega-9 content, makes it better suited as a biodiesel precursor. The presence of MUFAs and omega-9 fatty acids enhances the lipid profile for biodiesel production by improving the fuel's oxidative stability and lowering its cloud point, thus meeting more of the biodiesel industry standards.
4. Conclusion
This study demonstrates that pre-treatment with natural deep eutectic solvents (NaDES) can significantly enhance the efficiency of lipid extraction from Chlorella vulgaris. The application of NaDES disrupts the cell wall, facilitating easier access for solvents to interact with intracellular lipids. As a result, the total lipid yield from the biomass treated with
NaDES varies depending on the specific NaDES formulation and molar ratio used. The highest total lipid yield, amounting to 19.44% (w/w), was observed with the NaDES variation of choline chloride (ChCl) and glycerol at a 1:1 molar ratio. Gas chromatography-mass spectrometry (GC-MS) analysis further reveals variations in the fatty acid profiles of the extracted lipids, with a predominance of C16 (palmitic acid) and C18 (stearic and oleic acids) fatty acid chains. These findings underscore the potential of NaDES pre-treatment not only to improve lipid yield but also to influence the fatty acid composition, which is crucial for subsequent applications such as biodiesel production.
Acknowledgements
We would like to express our gratitude to the School of Life Sciences and Technology, Institut Teknologi Bandung for providing the essential laboratory equipment that made this research possible. Additionally, we extend our thanks to our research group members, Jonathan and Jeffry, for their invaluable assistance and support in the laboratory. Their contributions were instrumental to the success of this study.