
1 Pendahuluan
Secara in vivo, DNA polimerase mempunyai peranan yang sangat penting yaitu dalam proses replikasi dan perbaikan kesalahan DNA (repair). Fungsi utama aktivitas enzim tersebut adalah mengkatalisis penambahan unit mononukleotida dari deoksiribonukleosida 5'-trifosfat (dNTP) pada ujung 3'-OH bebas suatu untaian primer. Dalam reaksi tersebut diperlukan satu untai DNA yang digunakan sebagai cetakan (template) yang mengarahkan enzim dalam menyeleksi nukleotida yang masuk <sup>(1)</sup>.
Beberapa DNA Polimerase telah diidentifikasi dan dikarakterisasi baik dari eubakteria maupun archaea termofilik. Taq Pol I (\(Thermus\ aquaticus\\)Pol I) merupakan enzim polimerase yang pertama kali diisolasi dari eubakteria termofilik \(^{(2)}\). Umumnya DNA polimerase yang berasal dari genus ini tidak mempunyai aktivitas eksonuklease \(3^{\sim} \rightarrow 5^{\sim}\) yang merupakan aktivitas proofreading. Hal yang sama juga terdapat pada DNA Pol I dari genus Bacillus, seperti \(B.\ caldotenax\\)dan \(B.\ stearothermophylus\) \(^{(2)}\) \(^{(3)}\).
Enzim DNA Pol I dari B. stearothermophylus telah dikarakterisasi dan dimurnikan. Hasil karakterisasi diketahui bahwa enzim hasil pemurnian mempunyai suhu optimum \(65-72^{\circ}\)C dengan nilai kcat enzim tersebut 35 kali lebih tinggi dibanding enzim yang berasal dari KF (Klenow Fragmen) dan 4 kali lebih tinggi dibanding Taq Polimerase. Enzim ini juga mempunyai aktivitas spesifik yang lebih besar dibandingkan Taq dan KF Polimerase, dengan menggunakan substrat M13 dan calf thymus <sup>(4)</sup>. Hal yang sama juga terjadi pada prosesivitasnya yaitu 14 kali lebih tinggi dibanding KF Polimerase dan 2 kali lebih tinggi dibanding Taq Polimerase <sup>(4)</sup>. DNA Polimerase yang berasal dari archaea termofilik antara lain dari Pyrocoocus furiosus, Pyrodictum occultum,dan Sulfolabus solfaataricus, dikenal sebagai DNA Pol \(\alpha\) yang umumnya mempunyai aktivitas eksonuklease \(3^{\circ} \rightarrow 5^{\circ}\) namun tidak mempunyai aktivitas eksonuklease \(5^{\circ} \rightarrow 3^{\circ}\) <sup>(3)</sup>.
Gen penyandi DNA Pol I yang berasal dari bakteri termofilik isolat lokal telah diklon dan dikarakterisasi. Hasil penentuan urutan nukleotida diperoleh panjang gen adalah 2631 pb yang mengkode 876 asam amino <sup>(5)</sup>. Gen tersebut telah diekspresi secara heterolog di E.coli namun struktur - fungsinya belum diketahui <sup>(6)</sup>. Studi mutagenesis pada gen penyandi DNA Pol I telah dilakukan, antara lain mutasi tunggal pada residu asam amino Asp<sup>802</sup> dan gen mutannya (D802N) telah dikonstruksi dan diekspresikan secara heterolog di E.coli <sup>(7)</sup>. Tulisan ini melaporkan karakterisasi enzim mutan D802N DNA Pol I ITB-1 yang dibandingkan dengan enzim wild-type DNA Pol I ITB-1.
2 Metode
2.1 Peremajaan E.coli
E.coli GI724 yang membawa plasmid rekombinan (pITB-9A) dan (pITB-9MN) yang telah termutasi digoreskan dalam medium RMG-A (1 X M9 salts, 2% casamino acid, 0,5% glukosa, 1 mM MgCl<sub>2</sub>, 100 μg/ml ampisilin, dan 1,5% bacto-agar) langsung dari stok gliserol kemudian diinkubasi pada suhu 30°C selama 12-16 jam (sesuai manufaktur, Invitrogen).
2.2 Isolasi Enzim Wild-type dan Mutan D802N DNA Pol I ITB-1
Enzim wild-type dan mutan D802N DNA Pol I ITB-1 diperoleh dengan menumbuhkan masing-masing koloni tunggal E. coli yang membawa plasmid rekombinan pITB-9A dan pITB-9MN dalam medium RMG-A selama 16 jam dengan pengocokan 150 rpm pada 30°C (sesuai manufaktur, Invitrogen). Selanjutnya kultur sel dipindahkan ke dalam media induksi (0,2% Casamino Acids, 0,5% Glukosa, 1 mM MgCl<sub>2</sub>, Garam M9 1X, 100 µg/ml ampisilin) hingga OD<sub>550</sub> mencapai 0,1 dan inkubasi dilanjutkan hingga OD<sub>550</sub> mencapai 0,5. Selanjutnya ke dalam media tersebut ditambahkan triptofan hingga konsentrasinya 100 μg/ml, dan dilakukan pengocokan selama 3 jam pada 37°C. Pelet sel diperoleh dengan cara memisahkan sel dari media, yaitu dengan cara sentrifugasi pada 6000 x g selama 10 menit. 4°C. Pelet sel dicuci dengan bufer Tris-Cl (50 mM KCl; 10 mM Tris-Cl pH: 7,4; 0,1% Triton X-100; 2,5 mM MgCl<sub>2</sub>), kemudian disentrifugasi kembali pada 6000 x g selama 10 menit pada 4°C (pencucian dilakukan 2 kali) dan diperoleh pelet sel yang sudah bebas dari media. Ekstrak enzim kasar diperoleh dengan cara memecah pelet sel yang sudah bersih menggunakan ultrasonikator pada keadaan dingin 4°C. Debris sel dipisahkan dengan cara sentrifugasi dengan kecepatan 12000 x g selama 10 menit pada 4°C dan supernatannya merupakan ekstrak enzim kasar.
2.3 Penentuan Aktivitas Enzim Wild-type dan Mutan D802N DNA Pol I ITB-1
Penentuan aktivitas enzim wild-type dan mutan D802N dilakukan dengan metode radioaktif <sup>(8)</sup> yang didasarkan pada pengendapan DNA dengan TCA (Trichloroacetic Acid) yang merupakan jumlah <sup>32</sup>P-dATP yang terkorporasi pada DNA (calf thymus teraktivasi), endapan yang terbentuk diukur dengan Geiger Counter.
Reaksi enzim dilakukan sebagai berikut: 20 μl campuran yang mengandung 1,25 μg DNA calf thymus teraktivasi, 10 mM bufer Tris-Cl, dan 4 μl ekstrak enzim kasar wild-type dan mutan D802N DNA pol I ITB-1. Campuran reaksi
diinkubasi pada 65°C selama 15 menit dan reaksi dihentikan dengan menambahkan 0,5 M EDTA kemudian didinginkan pada 0°C. Selanjutnya sebanyak 5 µl campuran tersebut diteteskan pada kertas saring Whatman GF/C (dilakukan pada 2 kertas saring dan masing-masing dipipet 5 µl). Masing-masing kertas saring dikeringkan di bawah lampu Wolfram selama 5 menit, kertas saring pertama dicuci dengan 10 ml larutan 5 % TCA yang mengandung 20 mM natrium pirofosfat sambil digoyang (tahap pencucian ini dilakukan 2 kali), dan tahap akhir dicuci dengan etanol 75% kemudian dikeringkan kembali di bawah lampu Wolfram selama 10 menit. Kertas saring lainnya dibiarkan kering tanpa pencucian. Selanjutnya masing-masing kertas saring yang sudah kering dicacah dengan menggunakan Geiger Counter (9).
Aktivitas enzim didefinisikan sebagai unit aktivitas yang merupakan jumlah deoksinukleotida (pmol) yang terkorporasi per ml enzim x menit<sup>-1</sup> pada kondisi optimum.
2.4 Karakterisasi Enzim Wild-type dan Mutan D802N DNA Pol I ITB
Karakterisasi enzim meliputi: penentuan pH dan suhu optimum, penentuan kinetika enzim dengan menentukan nilai \(K_{m(dNTP)}\), \(V_{maks}\).
Penentuan pH dan suhu optimum dilakukan seperti pada percobaan 2.3 dengan melakukan variasi pH dan Suhu, sedangkan kinetika enzim dilakukan seperti percobaan di atas hanya dengan melakukan variasi konsentrasi substrat dNTP. Harga \(K_m\) dan \(V_{maks}\) ditentukan dengan persamaan LWB (Lineweaver-Burk).
2.5 Penentuan Kadar Protein
Konsentrasi protein ditentukan dengan menggunakan metode Bradford<sup>(10)</sup> dengan menggunakan BSA sebagai standar.
3 Hasil dan Pembahasan
3.1 Isolasi Enzim Wild-type dan Mutan D802N DNA Pol I ITB-1
Dalam penelitian ini enzim DNA polimerase diekspresi setelah kultur diinduksi dengan triptofan, dan selanjutnya diinkubasi selama 3 jam pada 37°C. Enzim wild-type dan mutan D802N diisolasi dari E.coli yang membawa plasmid rekombinan DNA pol I wild-type dan DNA pol mutan yang ditumbuhkan dalam 50 ml medium induksi yang mengandung ampisilin. Ekstrak enzim kasar diperoleh setelah sel dipecahkan dengan menggunakan ultrasonikator dengan frekuensi 30.000-40.000 kC selama 15 menit. Untuk mempertahankan agar
enzim yang diisolasi tidak terdenaturasi, pemecahan sel dilakukan dalam keadaan dingin yaitu pada 4o C dengan interval waktu 1 menit. Dari 50 ml kultur sel E.coli GI724 yang membawa plasmid rekombinan DNA pol I wildtype diperoleh berat sel 242,40 mg dengan total protein 9,95 mg. Sedangkan kultur sel DNA pol I mutan D802N diperoleh berat sel 232,90 mg dengan total protein 8,90 mg (Tabel 1). Analisis lebih lanjut dilakukan untuk menentukan aktivitas polimerase yaitu dengan mengukur jumlah cacahan (DNA yang terkorporasi) dari aktivitas enzim mutan D802N yang dibandingkan dengan enzim wild-type. Dalam penentuan ini digunakan DNA calf thymus teraktivasi sebagai (template-primer). Hasil analisis aktivitas spesifik (unit/mg protein) dari masing-masing enzim diperoleh 3,71 unit/mg protein untuk enzim wild-type dan 1,34 unit/mg untuk enzim mutan D802N. Data analisis menunjukkan terjadi penurunan aktivitas spesifik pada enzim mutan D802N yaitu sekitar 63,88 % (Tabel 1).
No Sampel Berat Sel (mg) Vol. (ml) Total Protein (mg) Aktivitas pmol/ml/men (103 ) Total Unit Akt. Spe (Unit/mg) 1 Wild-type 242,4 2,5 9,95 14,18 37,00 3,71 2 Mutan D802N 232,9 2,5 8,90 4,77 11,93 1,34
Tabel 1 Penentuan berat sel dan total protein enzim wild-type dan mutan D802N.
3.2 Karakterisasi Enzim Wild-type dan Mutan D802N DNA Pol I ITB-1
Untuk mengetahui lebih jauh pengaruh mutasi D802 menjadi N802, telah dilakukan karakterisasi terhadap enzim wild-type dan enzim mutan D802N. Karakterisasi meliputi penentuan pH dan suhu optimum, selain dari pada itu juga ditentukan kinetika enzimatiknya yang meliputi penentuan nilai Km dan Vmaks serta parameter kinetika lainnya seperti kcat dan kcat/Km.
3.2.1 pH dan Suhu Optimum Enzim Wild-type dan Mutan D802N DNA Pol I ITB-1
Penentuan pH dan suhu optimum enzim dilakukan berkaitan dengan sifat-sifat protein seperti kelarutan, kekuatan interaksi antara asam-asam amino, dan secara umum akan menyebabkan protein menjadi tidak stabil. Hasil analisis diperoleh nilai pH optimum (Gambar 1) dan suhu optimum (Gambar 2) untuk enzim wild-type dan enzim mutan D802N. Nilai pH dan suhu optimum enzim wild-type berturut-turut adalah 7,4 dan 65°C; Sedangkan enzim mutan D802N 9,0 dan 50°C. Pada keadaan optimum, enzim mempunyai aktivitas yang tinggi. Kondisi ekstrem seperti pH dan suhu tinggi (di atas kondisi optimumnya), enzim akan kehilangan aktivitas katalitiknya. Adanya penggantian gugus pada residu D menjadi N menyebabkan aktivitas enzim mutan turun bila dibandingkan dengan enzim wild-type (Tabel 1). Perubahan pH dapat menyebabkan ionisasi pada struktur primer dan struktur sekunder protein, juga menyebabkan perubahan interaksi antara satu asam amino dengan asam amino lainnya yang bertanggung jawab dalam pembentukkan struktur tiga dimensi.
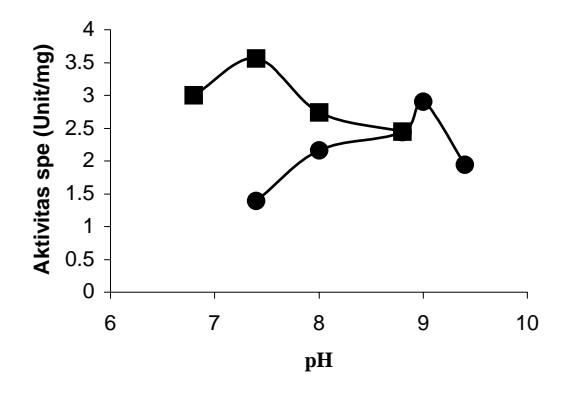
Gambar 1 Pengaruh pH terhadap aktivitas DNA Pol I ITB-1 wild-type dan mutan D802N. (Wild-type); (mutan D802N).
Berdasarkan hasil analisis menunjukkan bahwa mutasi asam amino D<sup>802</sup> meniadi N<sup>802</sup> menvebabkan terjadinya pergeseran nilai optimum baik terhadap pH maupun suhu enzim. Pada penentuan pH optimum terjadi kenaikan pH enzim mutan D802N, perubahan ini mungkin disebabkan karena asam amino sebelum dimutasi D<sup>802</sup> mempunyai rantai samping dengan gugus R bermutan negatif sedangkan setelah dilakukan mutasi menjadi N<sup>802</sup> dengan samping mengandung gugus R polar dan tidak bermuatan. Perubahan ini dapat dijelaskan pada keadaan keseimbangan protonik dari asam amino D dan N. Pada pH sekitar 6-8 asam amino D bermuatan negatif 1. Hasil percobaan menunjukkan bahwa enzim wild-type mempunyai aktivitas polimerase pada pH optimum 7,4. Sedangkan asam amino N pada kondisi yang sama tidak bermuatan (mempunyai muatan netto/bersih nol), sehingga untuk meningkatkan agar bermuatan negatif 1 (sama seperti pada D) maka asam amino tersebut harus berada pada pH basa. Hasil percobaan diperoleh aktivitas enzim mutan D802N mempunyai pH optimum 9,0, hal ini menyarankan bahwa muatan pada residu asam amino posisi 802 penting untuk aktivitasnya.
Hal yang sama terjadi pada penentuan suhu optimum enzim. Pada keadaan ini kecepatan reaksi enzim paling cepat. Kenaikan temperatur menyebabkan enzim terdenaturasi sehingga aktivitas katalitiknya berkurang. Dari data yang dihasilkan terlihat pada suhu 60°C terjadi penurunan aktivitas spesifik enzim mutan D802N sebesar 77% yaitu dari 4,23 unit/mg protein menjadi 1,0 unit/mg protein (Gambar 2).
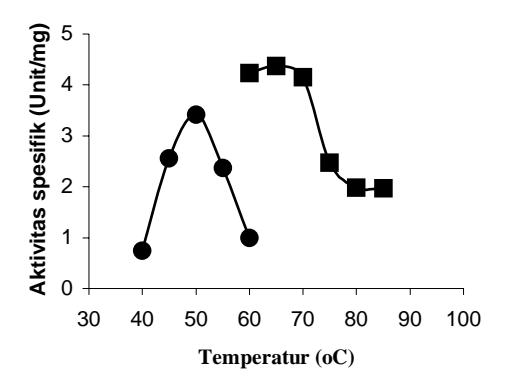
Gambar 2 Pengaruh suhu terhadap aktivitas DNA Pol I ITB-1 wild-type dan mutan D802N. (wild-type); (mutan D802N).
Berdasarkan hasil analisis menunjukkan adanya penurunan suhu optimum yaitu dari 65°C untuk enzim wild-type menjadi 50°C untuk enzim mutan D802N. Turunnya suhu optimum pada enzim mutan D802N hanya disebabkan oleh adanya mutasi D<sup>802</sup> menjadi N<sup>802</sup> sehingga sehingga kemungkinan perubahan tersebut mengakibatkan perubahan konformasi pada struktur protein mutan yang mengakibatkan enzim mutan D802N menjadi tidak stabil dan aktivitas enzim berkurang.
3.2.2 Kinetika Enzim Wild-type dan Mutan D802N DNA PolI ITB-1
Kinetika enzim wild-type dan enzim mutan D802N ditentukan dengan mengukur aktivitas masing-masing enzim pada variasi konsentrasi substrat dNTP dengan konsentrasi substrat lainnya yaitu DNA calf thymus dalam keadaan jenuh dan nilai \(K_{m(dNTP)}\) ditentukan dengan menggunakan kurva Lineweaver-Burk (Gambar 3). Nilai \(K_m\) menunjukkan afinitas enzim terhadap substrat dan spesifik untuk setiap enzim. Semakin kecil nilai \(K_m\) maka afinitasnya semakin besar. Hasil analisis diperoleh nilai \(K_{m(dNTP)}\) dan \(V_{maks}\) enzim wild-type berturut-turut adalah 0,95 \(\mu\)M dan 20,119 x 10<sup>3</sup> pmol/menit. Sedangkan nilai \(K_{m(dNTP)}\) dan \(V_{maks}\) enzim mutan D802N adalah 1,37 \(\mu\)M dan 15,737 x 10<sup>3</sup> pmol/menit (Gambar 3).
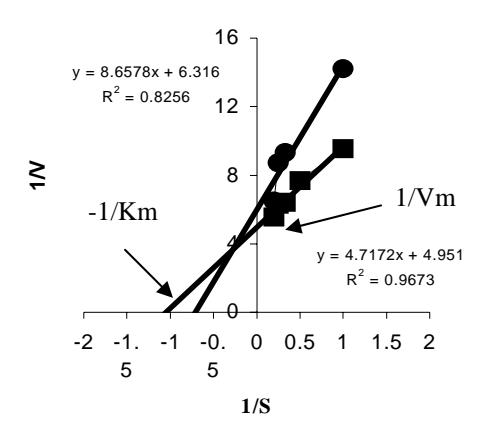
Gambar 3 Nilai Km(dNTP) dan Vmaks DNA Pol I ITB-1 wild-type dan mutan D802N. (wild-type); (mutan D802N).
Selain nilai Km dapat juga ditentukan nilai kcat enzim yang menggambarkan jumlah maksimum molekul substrat yang dikonversi menjadi produk (turn over number). Berdasarkan hasil percobaan, mutasi D802N juga memberikan pengaruh terhadap nilai kcat . Kenaikan nilai Km(dNTP) menyebabkan nilai kcat berkurang 13% bila dibandingkan enzim wild-type (Tabel 2). Parameter kinetika lainnya yaitu kcat/Km yang menggambarkan effisiensi katalitik. Tabel 2 menunjukkan terjadi penurunan nilai kcat/Km untuk enzim mutan D802N ± 2 kali dibanding enzim wild-type.
Perubahan nilai Km dan Vmaks diduga karena terjadinya perubahan pH dan suhu optimum enzim. Berdasarkan hasil-hasil yang diperoleh, menunjukkan bahwa perubahan pH dan suhu optimum pada enzim mutan D802N menyebabkan nilai Km meningkat 1,5 kali dibandingkan enzim wild-type. Peningkatan nilai menunjukkan penurunan affinitas enzim terhadap substrat dNTP demikian pula dengan pembentukan produk (dari data diperoleh nilai Vmaks menurun). Hasil penelitian yang dilakukan oleh Copeland(11) yaitu dengan melakukan mutasi pada daerah II (motif DFNSLYPSII) menunjukkan bahwa nilai Km enzim mutan meningkat bila dibandingkan enzim wild-type. Kelompok peneliti lain juga telah melaporkan hasil-hasil penelitiannya dengan melakukan mutasi terhadap residu asam amino yang mengandung gugus karboksilat, yaitu mutasi (D705E, D705A); (E710D, E710A); (D882E, D882A); dan (E883D, E883A) pada polimerase yang berasal dari Klenow Fragment (KF). Hasil penelitian menunjukkan bahwa terjadi penurunan terhadap aktivitas polimerase baik yang dimutasi menjadi E maupun A dengan penurunan diantara 30-70% (8).
| No | Sampel | kcat Menit-1 | Relatif kcat | kcat/Km M-1.menit-1 |
|---|---|---|---|---|
| 1 | Wild-type | 0,64 | 1,00 | \(6.7 \times 10^3\) |
| 2 | Mutan D802N | 0,56 | 0,87 | 4,1 x 103 |
Tabel 2 Parameter kinetika enzim wild-type dan enzim mutan D802N.
4 Kesimpulan
Berdasarkan hasil percobaan, enzim mutan D802N DNA Pol I ITB-1 mempunyai sifat berbeda dengan enzim wild-type. Enzim mutan D802N mempunyai pH dan suhu optimum pada 9,0 dan 50°C dengan aktivitas spesifik 1,34 unit/ mg protein; Sedangkan enzim wild-type mempunyai pH dan suhu optimum pada 7,4 dan 65°C dengan aktivitas spesifik 3,71 unit/ mg protein. Nilai \(K_{m(dNTP)}\) dan \(V_{maks}\) enzim mutan D802N 1.37 mM dan 15.832 x 10³ pmol/menit; Sedangkan nilai \(K_{m(dNTP)}\) dan \(V_{maks}\) enzim wild-type 0,95 mM dan 20.197 x 10³ pmol/menit. Hal ini menyarankan bahwa mutasi D<sup>802</sup> menjadi N<sup>802</sup> menyebabkan afinitas enzim terhadap substrat berkurang.
Ucapan Terima Kasih
Penelitian ini tidak dapat dilaksanakan dengan baik tanpa bantuan dana dari Riset Unggulan Terpadu X, Kementrian Riset dan Teknologi.
Daftar Pustaka
- 1. Joyce, C.M., & Steitz, T.A., Function And Structure Relationships In DNA Polymerases, Annu. Rev. Biochem, 63, 773 822 (1994).
- Blondal, T., Thorbjarnardottir, S.H., Kieleczawa, J., Hjorrleifsdottir, S., Kristjansson, J.K., Einarsson, J.M., Eggertsson, G. (2001). Cloning, Sequence Analysis and Functional Characterization of DNA Polymerase I from the Thermophilic Eubacterium Rhodothermus marinus. Biotechnol. Appl. Biochem. 34:37 – 45.
- 3. Pisani, F.M., Felice, M.D., Manco, G., & Rossi, M., Domain Organization and Biochemical Features of Sulfolobus solfataricus DNA Polymerase, Extremophile, 2, 171 177 (1998).
- 4. Kiefer, J.R., Mao, C., Hansen, C.J., Basehore, S.L., Hogrefe, H.H., Braman, J.C., & Beese, L.S., Crystal Structure of a Thermostable Bacillus DNA Polymerase I Large Fragment at 2.1 A<sup>o</sup> Resolutio, Structure, 5, 95-108 (1997).
- 5. Akhmaloka, Pramono H., Tika I.N., Suharto A., Retnoningrum D.S., Sindumarta M., Padmawinata K., dan Oei B.L., (2000b). Eksplorasi potensi bakteri termofilik isolat lokal: Studi kasus DNA polimerase termostabil, Prosiding Seminar Hasil Penelitian dan Pengembangan Bioteknologi III, Cibinong 7-9 Maret 2000: 31-39.
- 6. Pramono H., Kloning dan Ekspresi di Escherichia coli Gen DNA Polimerase I Bakteri Termofilik Isolat Cimanggu. Disertasi Doktor, Departemen Kimia, Institut Teknologi Bandung (2004).
- 7. Ambarsari, L., Pramono, H., Madayanti,F., Moeis, R.M., & Akhmaloka, Konstruksi dan Overekspresi Gen Penyandi DNA Polimerase I Mutan dari Bakteri Termofilik Isolat Lokal, J.Hayati, 11, 115-120 (2004).
- 8. Gangurde, R., N., Kaushik, Singh, K., & Modak, M.J., A Carboxylate Triad Is Essential for the Polymerase Activity of Escherichia coli DNA Polymerase I (Klenow Fragment), J. Biol. Chem, 275, 19685 – 19692 (2000).
- 9. Sambrook, J., Russell, D.W., Molecular Cloning: A Laboratory Manual, Ed., ke-3, Cold Spring Harbor Laboratory Press, New York (2001).
- 10. Bradford, M.M., A Rapid and Sensitive Method for the Quntitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding, Anal. Biochem., 72, 248-254 (1976).
- 11. Copeland, C., Dong, Q., Wang, T.S.F., Rational for Mutagenesis of DNA Polymerase Active Sites: DNA Polymerase α, Methods in Enzymology, 262, 294-303 (1995).