
1 Introduction
Polycyclic aromatic hydrocarbons (PAHs) are highly toxic organic pollutants which are widely distributed in terrestrial and aquatic environments. They consist of benzene rings which have two or more fused aromatic rings in linear, angular, or clustered alignment and are mainly formed as products of the incomplete combustion of fossil fuels. Until now, research on the microbial degradation of PAHs has been demonstrated that microorganisms such as bacteria, fungi and alga posses catabolic abilities that may be utilized for the remediation of PAH-contaminated soil and water. Recently, bioremediation has been shown to be effective in remediating environment contaminated with low molecular weight PAHs [1,2].
However, the high molecular weights PAHs are generally recalcitrant to microbial attack [3,4]. Although the lack of microbial activity towards high
molecular weight PAHs may be attributed to site specific environmental factors, such as bioavailability of the contaminant, nutrients, redox potential, etc., the limiting factor may be the scarcity of microorganisms capable degrading the more highly condensed compounds. For bioremediation to be an effective tool for the clean up of PAHs in environment, a greater understanding of the process involved, and those are limit the degradation of high molecular weight PAHs, is required.
Laccases (EC 1.10.3.2.) belong to the group of polyphenoloxidases and are usually considered to be enzymes able to oxidize aromatic compounds with a phenolic functional group. The classical action of laccases-oxidation of substrate by transferring electrons to oxygen in one-electron steps - results in the polymerization of phenols and/or the forming of quinones. Laccases can be applied extensively in many fields, including waste detoxification and textile dye transformation due to their low substrate specificity [5].
Recently, some researcher reported an oxidation of the 3-ring polycyclic aromatic hydrocarbon (PAH) anthracene by laccase of white rot fungi [6]. The anthracene removed was stoichiometrically oxidized to 9,10-anthraquinone. The reaction was enhanced significantly by adding of ABTS or 1 hydroxybenzotriazole (HBT). Analogously, oxidation of white rot fungi by laccase of white rot fungi with ABTS as the mediator compound was reported [6,7]. Our study presented here provides results on the degradation of 3 different rings PAH with the laccase of white rot fungus Polyporus sp. S133. We also describe the effect of a mediator on this reaction and the metabolites produced by the oxidation of PAH.
2 Experiments/Methods
2.1 Screening Microorganism
Screening of fungi was conducted by collecting soil, swamp, and sludge contaminated with petroleum and cultured in malt extract agar medium to grow the microorganism. The fastest fungi that grow in agar medium containing chrysene as test contaminant was stored in 4°C. The strain was maintained on malt extract agar (2% (w/v) malt extract, 2% (w/v) glucose, 0.1% (w/v) polypeptone, and 1.5% (w/v) agar) in a plastic Petri dish.
2.2 Chemicals
Phenanthrene, chrysene and benzo[a]pyrene (BaP) was purchased from Tokyo Chemical Industry Co. Ltd. (Tokyo, Japan). Malt extract and polypeptone were purchased from Difco (Detroit, USA). Thin layer chromatography (TLC) aluminium sheets (Silica gel 60 F<sub>254</sub>, 20x20cm) were obtained from Merk (Darmstadt, Germany). The silica gel used for column chromatography (wakogel S-1), and all other chemicals were purchased from Wako Pure Chemical Industry Co. Ltd. (Osaka, Japan).
2.3 Laccase Production
Time course of laccase production by Polyporus sp. S133 was studied in 100 ml malt extract medium at 30°C at static condition. Laccase activity (as mentioned in following section) was measured in crude cell extract of Polyporus sp. S133 cells grown at different time intervals. For higher laccase production, 10% inoculum of 12 h (A 660 nm) grown Polyporus sp. S133 was inoculated in 3 l nutrient medium and incubated 12 h at 30°C. Cells were collected by centrifugation at 8000 rpm for 15 min and suspended (150 mg ml<sup>-1</sup>) in 50mM sodium phosphate buffer (pH 7.0) containing 5 mg ml<sup>-1</sup> lysozyme. Cells were further incubated at 37°C for 45 min in water bath and then disrupted by sonication. This cell free extract was solubilized in cholic acid (0.33 mg mgprotein<sup>-1</sup>) on magnetic stirrer at 4°C for 30 min. The cell lysate obtained was centrifuged twice at 15,000 rpm for 30 min at 4°C and the clear supernatant used immediately or stored at -20°C until its use to purify laccase.
2.4 Purification of Laccase
The supernatant containing laccase activity 0.04 U (mg protein)<sup>-1</sup> min<sup>-1</sup> was heated at 60°C for 10 min and centrifuged at 8000 rpm for 20 min. The clear supernatant obtained after centrifugation was loaded on a DEAE cellulose fast flow column (15mm×120mm). The column was washed with the buffer by two times of the column volume and the enzyme was eluted with a linear gradient of 0–1M NaCl. Fractions containing laccase activity were pooled and dialyzed against 1mM sodium phosphate buffer (pH 6.0). The dialyzed sample was concentrated (1–2 ml) by ultrafiltration and loaded on Biogel column (10mm×500mm) equilibrated with 50mM sodium phosphate buffer (pH 6.0). The protein elution was carried with the same buffer at 6 ml h<sup>-1</sup> flow rate. Fractions containing laccase activity were pooled and stored at –20°C until use.
2.5 Protein Determination and Enzyme Activity
The protein concentration of each fraction was monitored by absorbance at 280nm or Lowry methods with bovine serum albumin as a standard [8]. Laccase
activity was determined at 30 °C by measuring increase in optical density at 420nm in a reaction mixture of 2 ml containing 0.66mM ABTS in 0.1M acetate buffer (pH 4.9) and \(100\mu l\) enzyme [9]. One unit of enzyme activity was defined as a change in absorbance unit min<sup>-1</sup> mgprotein<sup>-1</sup>.
2.6 Degradation of PAHs by Purified Laccase
Some PAHs were used in this study. The degradation reaction was carried out at 30°C for 24 h in 2 ml reactions mixture containing 100 mg l<sup>-1</sup> dye prepared in 50mM acetate buffer (pH 4.8) and 0.5 U ml<sup>-1</sup> purified laccase. Control containing heat-denatured enzyme was used to measure degradation of PAHs at different time interval. The above degradation samples were centrifuged at 1000 rpm for 1 min and laccase degraded metabolites were extracted by adding twice equal volume of ethyl acetate. After drying over anhydrous Na<sub>2</sub>SO<sub>4</sub>, the biodegraded metabolites were characterized by Gas Chromatography-Mass Spectrometry (GC-MS) and compared with standard PAHs. Gas Chromatography-Mass Spectrometry (GC-MS, Shimadzu QP-5050). GC-MS was performed with a column 30 m in length and 0.25 mm in diameter, and a helium pressure of 100 kPa. The temperature was initially 80°C, held for 2 minutes, raised from 80°C to 200°C at 20°C min<sup>-1</sup>, then to 260°C at 7.5°C min<sup>-1</sup>, and held for 4 minutes. The flow rate was 1.5 ml min<sup>-1</sup>, interface temperature was 260°C, and injection volume was 1µl.
3 Results and Discussion
A total of 10 samples of soil were inoculated on a malt extract agar medium containing chrysene. Among the 10 samples isolated, 20 fungi were selected for further screening by the method described above. Six fungi that grew well on the agar medium were selected among the 20. Other isolates were discarded based on their comparatively poor growth on agar medium containing chrysene. One fungus, named S133, was found to be most capable of degrading chrysene. S133 has a cap with an indented cell similar to a honeycomb white in color. Its stem is short, unequal, flared upward, and bent with the cap. Its spore is white, grows in groups of several specimens or in tuft of two or three individuals sometime with the caps welded together, and has clamp connection between hyphae [10,11]. Based on these macroscopic morphological characteristics, S133 was identified as Polyporus sp. S133.
PAH selected for the experiments represent compounds of great environmental significance. All experiments were performed in a buffered solution containing 2.5% acetone to increase the solubility of substrates. Recovery of PAH was generally in the range of 85–100%. All results were calculated and presented as percent of the control samples processed with heat-inactivated enzyme. Laccase of Polyporus sp. S133 was able to oxidize most of the PAH tested (Figure 1). Phenanthrene was removed to 89% from the reaction mixture during the 72 h incubation, followed by chrysene and benzo[a]pyrene which were oxidized to 66% and 55%, respectively. The oxidation of chrysene and benzo[a]pyrene was reported to be considerably enhanced by the addition of mediator compounds [12,13].
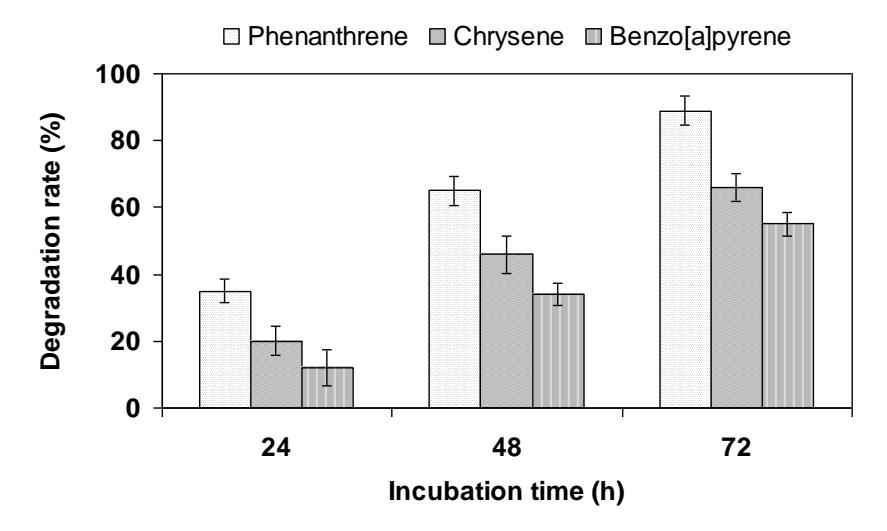
Figure 1 Oxidation (% of control) of polycyclic aromatic hydrocarbons (PAH) by laccase of Polyporus sp. S133.
1-Hydroxybenzotriazole displayed an especially high increase in the reaction rate. Application of this compound to the oxidation of PAH in our present study also showed a strong influence (Figure 2). Phenantrene were almost completely removed from the reaction mixture (98%). Oxidation of chrysene and benzo[a]pyrene increased 12 and 10% with the mediator to 78 and 65%, respectively, in the presence of HBT. Several lines of studies revealed that catalytic oxidation by laccases can be enhanced with the coexistence of redox mediators such as HBT [14]. Of the laccase systems tested, the laccase with HBT system was found to be the most effective [15]. It has been reported that the stability of the oxidized form of a mediator might play a key role, in addition to its redox potential [16]. It appeared that the oxidized form of HBT was more stable than that of the other mediators tested in the present system.
The TLC analyses were initially performed to indicate the presence of different metabolites in the degradative pathway of the PAHs culture. GC-MS was carried out to conclusively prove the presence of these metabolites. Degradation products were detected during the degradation of PAHs by Polyporus sp. S133.
These compounds were formed during the reaction and were identified by GC-MS with comparison of their mass spectra with synthetic and synthesized compounds after separation by column chromatography. These compounds were also the end product and were not further degraded in the system without the mediator during 72 h incubation.
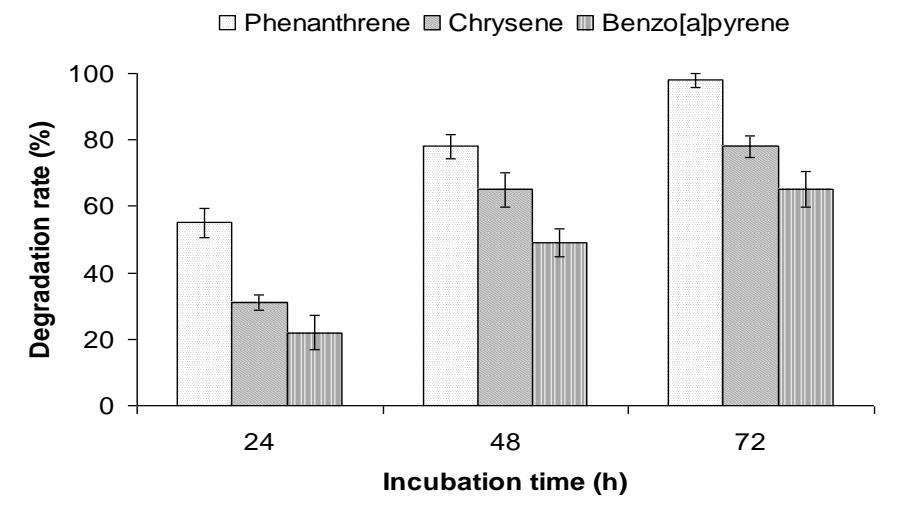
Figure 2 Oxidation (% of control) of polycyclic aromatic hydrocarbons (PAH) by laccase of Polyporus sp. S133 in the presence of 1-hydroxybenzotriazole (HBT).
Degradation product of phenantrene having an \(R_{\rm f}\) value of 0.44, similar to that of synthetic 9,10-phenanthrenequinone. The spectrum of this compound (m/z 208, M<sup>+</sup>), which had a retention time (\(t_{\rm R}\)) of 13.4 min, is shown in Figure 3A. The GC retention time, MS properties of the M<sup>+</sup> at m/z 208, and the significant fragment ions at m/z 152 and 180 (M<sup>+</sup>–28), corresponding to the respective sequential losses of –CO, were identical to those of synthetic 9,10-phenanthrenequinone.
Degradation product of chrysene (m/z 258, \(M^+\)) having an \(R_f\) value of 0.35, similar to that of synthesized 6,7-chrysenequinone. The spectrum of this compound which had a retention time (\(t_R\)) of 3.1 minutes is shown in Figure 3B. The GC retention time, MS properties of the \(M^+\) at m/z 258, and fragment ions at m/z 230 (\(M^+\)-28), corresponding to the respective sequential losses of -CO, were identical to those of synthesized 6,7-chrysenequinone.
Degradation product of benzo[a]pirene having an R<sub>f</sub> value of 0.5, similar to that of synthesized benzo[a]pirene-6,12-quinone. The spectrum of this compound
(m/z 258, \(M^+\)) that had a retention time (t<sub>R</sub>) 19.6 min is shown in Fig. 3C. The GC retention time, MS properties of the \(M^+\) at m/z 258, and the significant fragment ions at m/z 282, 267 and 230 (\(M^+\)–28), corresponding to the respective sequential losses of –CO, were identical to those of synthesized benzo[a]pirene-6,12-quinone.
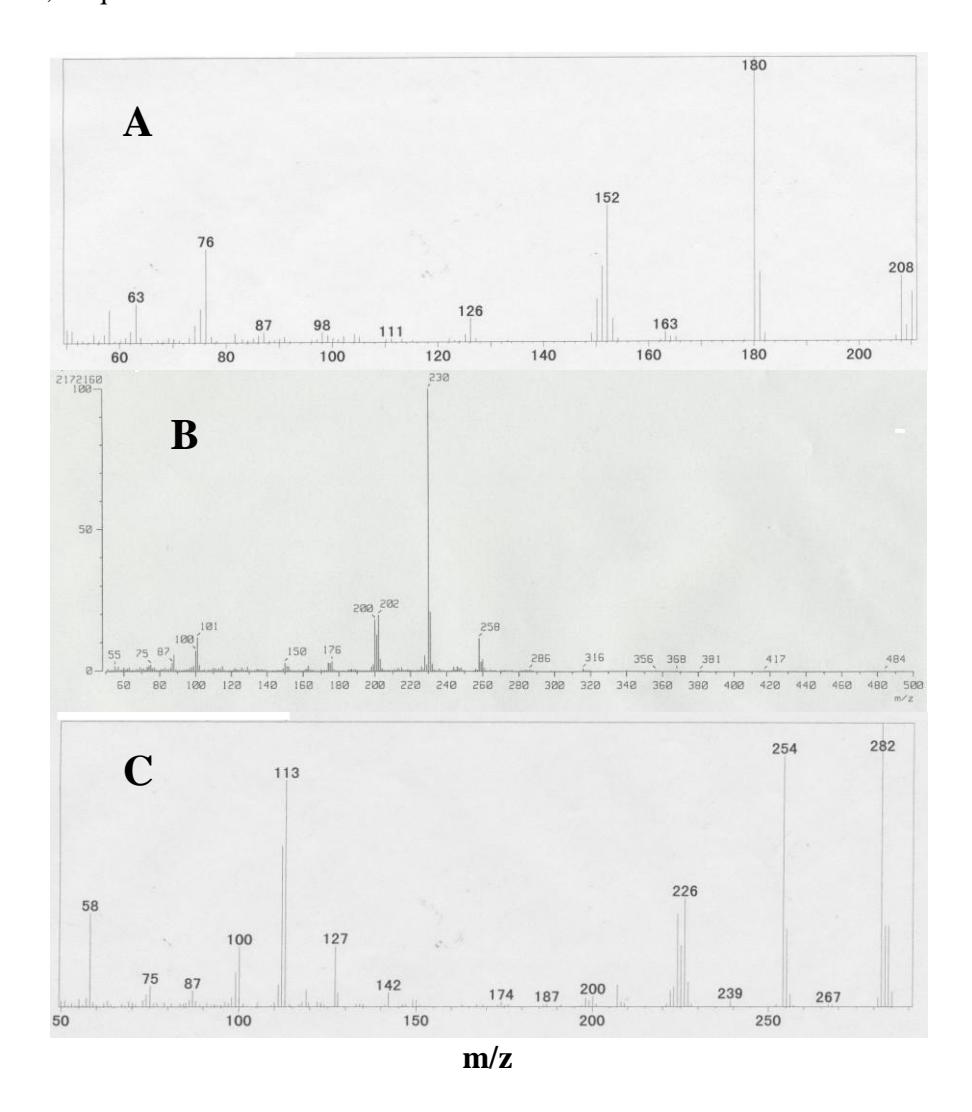
Figure 3 Mass Spectra of oxidation products of phenanthrene (a), chrysene (b) and benzo[a]pyrene c).
O O Chrysene 6,7-Chrysenequinone O O Benzo[a]pyrene-6,12-quinone Benzo[a]pyrene O O Phenanthrene 9,10-phenanthrenequinone
Figure 4 Pathways for the degradation of PAHs by Polyporus sp. S133.
These results showed that 9,10-, 6,7- and 6,12-oxidation and ring cleavage to give quinone structure was the major fate of PAHs in ligninolytic Polyporus sp. S133 (Figure 4). PAHs metabolism in ligninolytic Polyporus sp. S133 differs from the pathway employed by most bacteria, which cleave this PAH between C3 and C4 [17], and also differs from the process in nonligninolytic fungi and other eukaryotes, which are incapable of PAH ring fission [17,18]. Even though the obtained extracts were colored as in the case of PAHs, only very small amounts of products were detected in GC-MS analysis; however, a low but significant lifting of the background by GC-MS analysis corresponding to the degree of the removal of these PAH was observed. Analysis of the samples by size exclusion chromatography (SEC) confirmed the supposition of the polymeric nature of the products. Laccase did not polymerize HBT which was obvious from the results obtained with most PAH where no polymeric material was found and from control samples without PAH. Due to multiple oxidation products (formed only to a slight degree) and unknown structures and absorption coefficients of polymeric products, no attempt was made to quantify oxidation products of all single PAH. Quinones and hydroxy derivatives were reported to be common end products in the biodegradation of PAH by fungi [19]. Certain ring fission reactions in the subsequent steps of metabolization make a complete mineralization of PAH to carbon dioxide and water possible.
Acknowledgements
A part of this research was financially supported by a Grant-in-Aid for Scientific Research from Ministry of Education, Culture, Sports, Science and Technology of Japan (No. 20580178), which is greatfully acknowledged.