
1 Introduction
The potato is one of the world's major staple foods, together with rice, wheat and maize. In Indonesia, the potato is known as an important commodity that is widely cultivated by farmers, especially in the highlands [1]. Kuntjoro [2] states that there are several constraints in the production of potato crops in Indonesia, including (1) the poor quality and quantity of potato seedlings, which is a major concern in Indonesian's potato production; (2) the conventional cultivation techniques employed; (3) topographic factors, where the availability of terrains with altitude and temperature appropriate for planting potatoes is very limited; and (4) the tropical climate that is an optimum condition for the proliferation of pests and diseases.
Using conventional techniques, the establishment of large quantities of highquality potato seedlings is rather difficult. To overcome this problem several researchers have carried out studies on potato propagation by tissue culture. One possibility is through somatic embryogenesis. Somatic embryogenesis is a process where somatic cells evolve to form a new plant through specific stages of embryonic development without gametes fusion. This method has been used for plant propagation as well as to speed up and increase the success of genetic transformation [3].
The success of somatic embryogenesis is affected by several factors: the source of the explant; media composition, including the combination of growth regulators; and the physical conditions of the culture, including light, temperature, pH and aseptic conditions [4]. Plant growth regulators (PGR) have an important role in determining the success of somatic embryogenesis. Somatic embryogenesis is commonly induced by using a single application of growth regulator, usually auxin (2,4-D). In some plants, however, somatic embryos can also be induced using a combination of auxin and cytokinin, i.e. BAP, kinetin, zeatin and 2-ip [1,5]. The choice of explant is important for somatic embryo induction, because the initiation of embryogenesis occurs only in responsive or competent cells [6]. The pattern of the developmental response is influenced by the stage of development: the embryogenic potential continuously decreases during plant ontogenesis and is species-dependent as well [7]. Embryonic tissue has the highest embryogenic potential. It declines in hypocotyl, petiole, leaf and roots [8]. Juvenile tissue, therefore, has a higher embryogenic potential than mature tissue [9]. Seedlings of carrot or alfalfa have a wide expression of embryogenic potential and any organ of their seedlings can be induced to develop somatic embryos [8]. In potatoes, somatic embryos have been successfully developed using in vitro leaf explants [4].
Somatic embryos can be used to support a breeding program to generate highquality potatoes. This research was conducted to evaluate the type of potato explants with the highest potential to produce somatic embryos; to evaluate the ability of different combinations of growth regulator (2,4-D and BAP) at various concentrations to induce potato somatic embryos; and to evaluate the ability of BAP to act as growth regulator for the potato somatic embryo maturation process.
2 Materials and Method
2.1 Callus Induction and Proliferation
In this study, young leaf and stem nodal sections from sprouting tubers of potato cv. Granola were used as sources of explants. These explants were then cultured in callus induction medium consisting of MS basal media [10] supplemented with 9 μM 2,4-D and 4.5 μM BAP for the stem nodal section explants and 9 μM 2,4-D and 1 μM BAP for the young-leaf explants. These media contained 30 g/L sucrose and were solidified with 2.5 g/L gelrite. The cultures were maintained at room temperature (18-21 ºC) in a 16/8 hour light/dark photoperiod. Subsequently, calli were subcultured every 2 weeks.
2.2 Somatic Embryo Induction
Friable and yellowish calli that developed from both types of explants were transferred into somatic embryo induction media, which consisted of MS basal media [10] supplemented with various concentrations of 2,4-D (1-10μM) and BAP (0.5-10μM) (Table 1). The cultures were agitated at 100 rpm and maintained at room temperature (18-21ºC) with a 16/8 hour light/dark photoperiod. The cultures were observed and sub-cultured every 2 weeks.
Table 1 Combination of growth regulators 2,4-D and BAP in somatic embryo induction media.
| 2,4-D (μM) | BAP (μM) | |||
|---|---|---|---|---|
| 10 | 5.0 | 1.0 | 0.5 | |
| 1.0 | A | B | C | D |
| 2.5 | E | F | G | H |
| 5.0 | I | J | K | L |
| 10 | M | N | O | P |
2.3 Somatic Embryo Development
The somatic embryos that developed on the somatic embryo induction medium were transferred into somatic embryo development media containing MS basal media supplemented with 1μM and 5μM BAP or MS basal media [10] without BAP. Sucrose (30 g/L) and activated charcoal (0.1 g/L) were added into the media. The culture was then incubated on a shaker (100 rpm) at room temperature (18-21 ºC) with a 16/8 hour light/dark photoperiod. The cultures were observed and sub-cultured every 2 weeks. Somatic embryo development was observed using an Olympus inverted microscope with 100x magnification.
3 Results and Discussion
3.1 Callus Induction of Potato (Solanum tuberosum L.)
Compact calli were the first to develop from both explants (Figure 1). These were formed after 4 weeks of culture. After sub-culturing 2-3 times, their appearance changed from compact (Figure 1A & 1B) to friable and yellowish
(Figure 1E & 1F). The growth of the stem nodal-derived calli was faster than that of the calli from leaf explants.
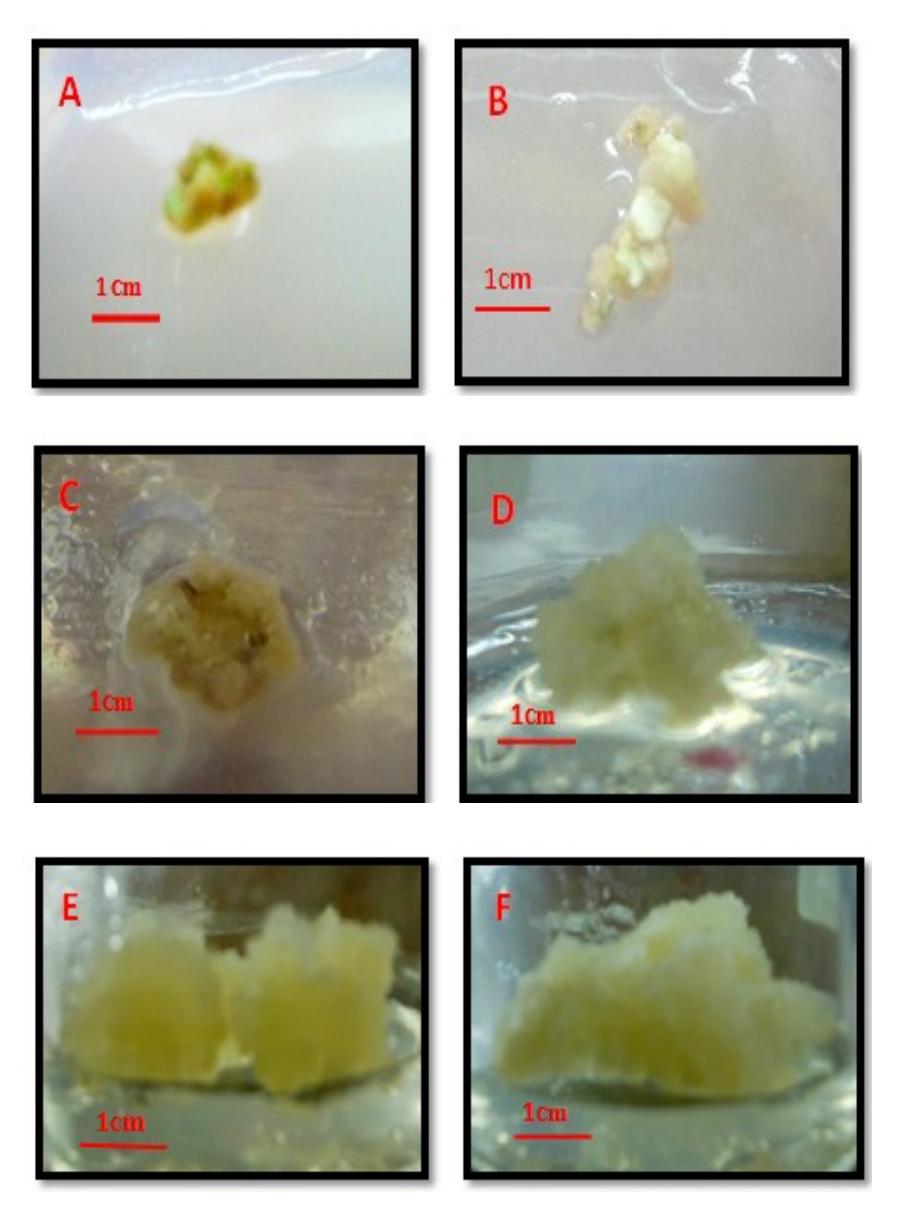
Figure 1 Calli from leaf explants (A, C, E) and stem nodal section explants (B, D, F) at 4, 8 and 16 weeks after culture.
The regeneration ability of callus depends on its structure. Purnamaningsih [11] states that friable, soft and yellowish callus usually has a higher regeneration
ability to form somatic embryos than compact callus. This form of callus is highly dependent on the type and concentration of PGR used. In this study, 2,4- D (auxin) and BAP (cytokinin) were able to induce calli from leaf explants and stem nodal section explants of potato plants. The calli were easily induced from the leaf explants in MS basal medium supplemented with 2,4-D 9 μM + BAP 4.5 μM, whereas the calli from the stem nodal section explants were induced in medium containing 2,4-D 9 μM + BAP 1 μM. The use of strong activity auxin (2,4-D) combined with generally low concentrations of cytokinin can induce friable callus [12]. In the present research, the combination of growth regulators used was probably able to balance the endogenous hormone produced by the explants for callus induction.
3.2 Somatic Embryo Induction and Development of Potato (Solanum tuberosum L.)
The friable calli were transferred to liquid medium for embryogenic callus induction. Two weeks after the initiation, embryogenic cells were already induced. The embryogenic cells could be easily distinguished from nonembryogenic cells (Figure 2). The embryogenic cells were mostly smaller, round-shaped with a dense cytoplasm and divided actively, while the nonembryogenic cells were mostly elongated and vacuolated, with less ability of cell division. The non-embryogenic cells were usually turned brown and eventually died. The embryogenic cells grew further, divided and developed to form somatic embryos.
Figure 2 Cell types found in cell suspension cultures of potato (Solanum tuberosum L.): (A) non-embryogenic cells (arrow) are large and elongated; (B) embryogenic cells (arrowhead) are smaller and divide actively.
Both explants used in this study were able to form embryogenic cells and somatic embryos (Figure 3 and Figure 4). Figure 3 and 4 show that the stem nodal-derived calli produced a higher amount of somatic embryos (in globular and heart stage) than the leaf explants, which may be due to stem nodal section explants being more responsive than leaf explants. According to Maillot, et al. [13], the meristematic cells in the lateral buds of stem nodal section explants have a higher embryogenic competence than leaf explants, therefore meristematic tissue is more sensitive to being induced into embryogenic cells compared with other somatic cells.
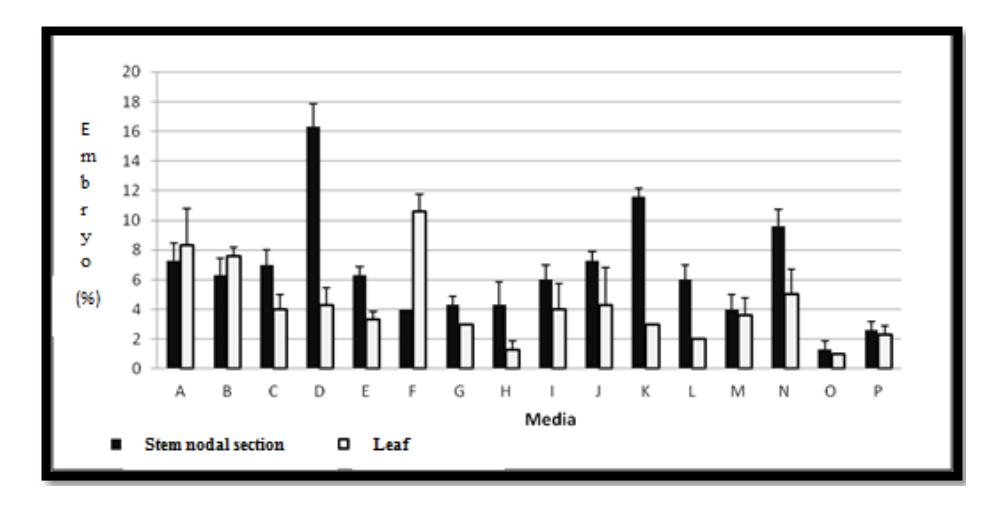
Figure 3 Percentage of globular stage somatic embryos from stem nodal section explants (■) and leaf explants (□) induced in different induction media.
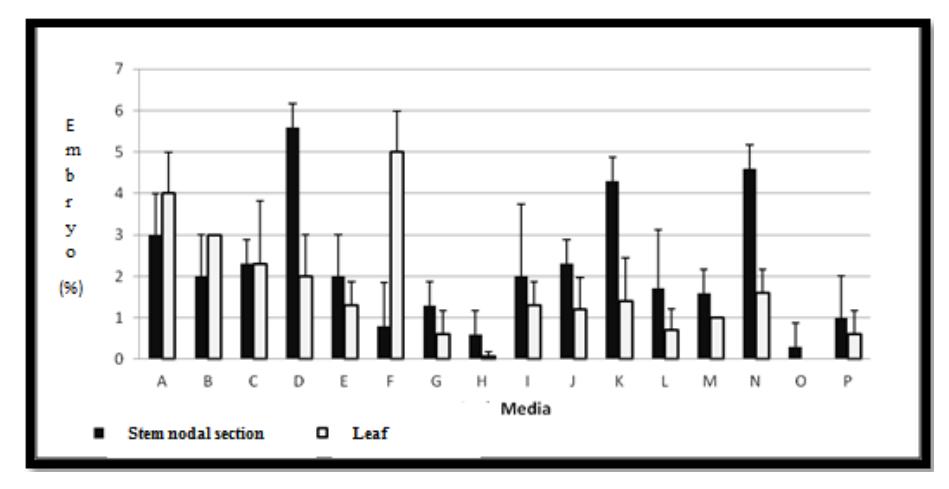
Figure 4 Percentage heart stage somatic embryos from stem nodal section explants (■) and leaf explants (□) induced in different induction media.
From the leaf explants, somatic embryos in globular and heart stage were mostly formed in the media containing 2.5 μM 2,4-D and 5 μM BAP; 1.0 μM 2,4-D and 10 μM BAP or 1.0 μM 2,4-D and 5.0 μM BAP. Meanwhile, from the stem nodal explants, induction of globular and heart stage somatic embryos was successfully achieved mostly in the media containing1.0 μM 2,4-D and 0.5 μM BAP; 5 μM 2,4-D and 1.0 μM BAP or 10 μM 2,4-D and 5.0μ M BAP. Based on these results, leaf-derived calli can be induced to form globular and heart stage somatic embryos in higher concentrations of BAP, whereas stem nodal-derived calli form somatic embryos in higher concentrations of 2,4-D. The difference is probably due to the higher concentration of auxin in leaf explants compared to stem nodal section explants. According to George, et al. [14], the highest content of auxin in plants is found in young leaf and shoots. Thus, the addition of exogenous auxin may no longer influence or can even inhibit growth. Kiyosue, et al. [15] state that the auxin concentration needed for somatic embryo induction is much higher than for normal cell growth. The importance of auxin for somatic embryo induction is also stated by Philips, et al. [16]: a high concentration of auxin is necessary for the induction of somatic embryogenesis.
3.3 Somatic Embryo Maturation of Potatoes (Solanum tuberosum L.)
In this study, somatic cells from both explants were capable to reach the torpedo stage of the somatic embryo through several changes in morphology. Initially, the potent somatic cell divides to produce a group of meristematic cells, called the proembryo. These meristematic cells continue to divide and subsequently give rise to the globular, heart shape and torpedo stages of the embryo (Figure 5).
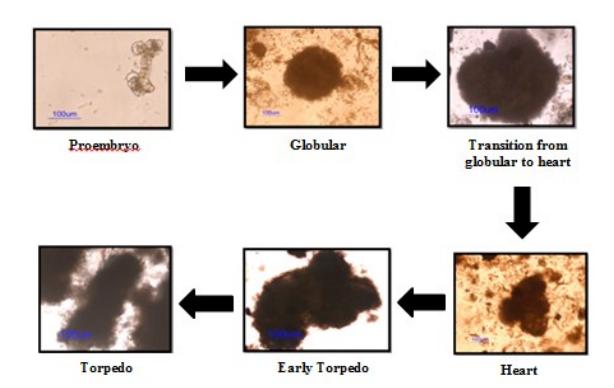
Figure 5 Development stages of somatic embryo of potato (Solanum tuberosum L.).
After 10 weeks in somatic embryo induction medium, the highest number and the fastest development of somatic embryos was obtained from the somatic embryos developed initially in media containing 2.5 μM2,4-D and 5.0μM BAP for the leaf explants and 10 μM 2,4-D and 5 μM BAP for the stem nodal section explants. Based on these results, somatic embryos from these two media were then selected for somatic embryo maturation by transferring them into somatic embryo maturation medium, which consisted of MS basal media supplemented with or without BAP. The development of the somatic embryo maturation process for each explant can be seen in Figure 6.
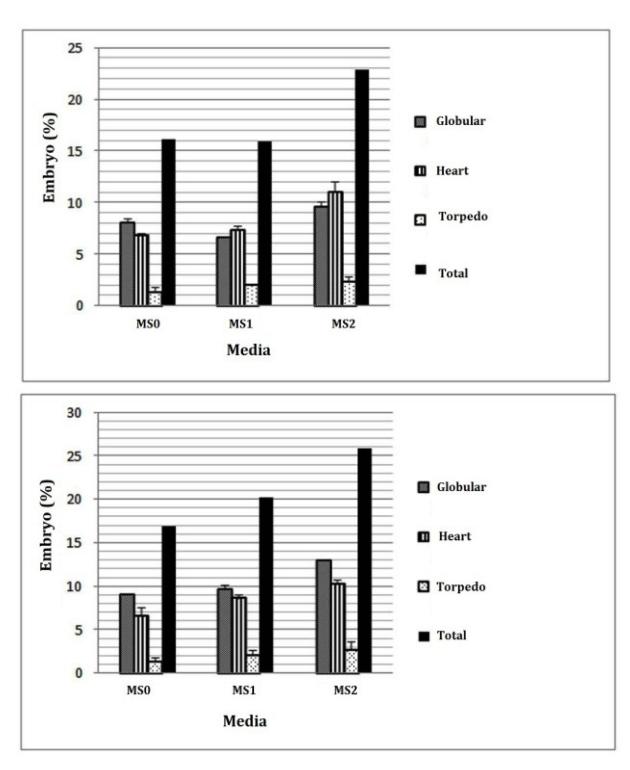
Figure 6 Development of somatic embryos derived from leaf explants (left) and stem nodal section explants (right) in maturation medium.
Figure 6 shows that the media containing 5 μM BAP was best for the development of somatic embryos. This suggests that cytokinin plays an important role in potato somatic embryo maturation. Priyono and Danimihardja [17] also note that the rate of the somatic embryo maturation process is influenced by the concentration level of cytokinin used, as cytokinin activates cell division (cytokinesis) and plant morphogenesis [1].
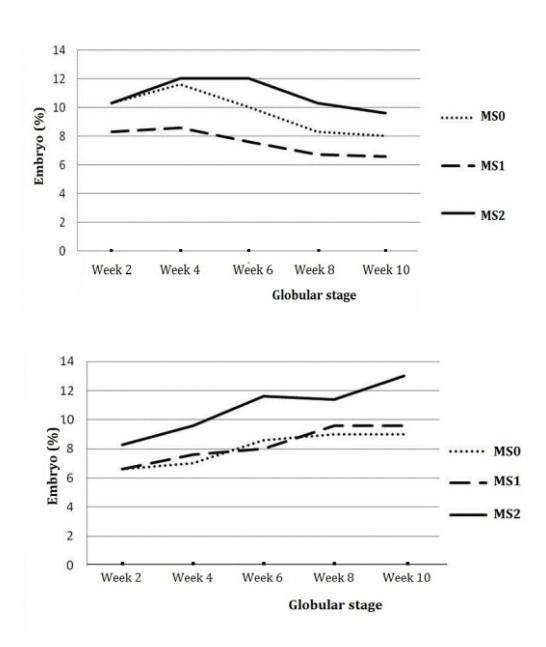
Figure 7 Development of globular stage somatic embryos derived from leaf explants (left) and stem nodal section explants (right) in maturation medium.
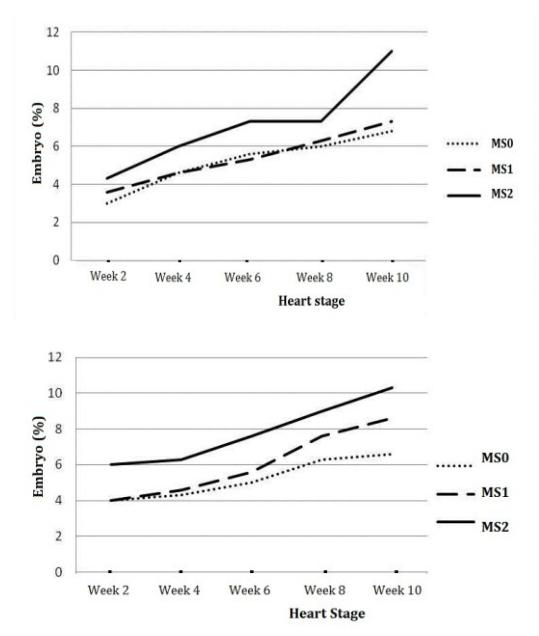
Figure 8 Development of heart stage somatic embryos derived from leaf explants (left) and stem nodal section explants (right) in maturation medium.
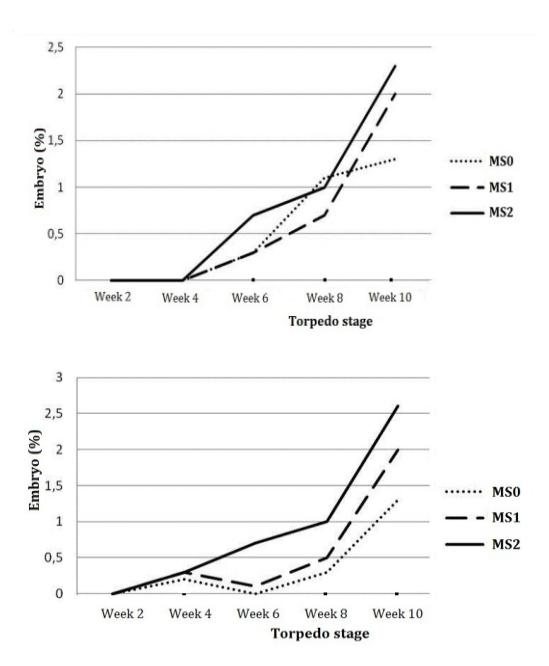
Figure 9 Development of torpedo stage somatic embryos derived from leaf explants (left) and stem nodal section explants (right) in maturation medium.
From Figure 7 it can be seen that the number of globular stage somatic embryos derived from the leaf explants increased from week 2 and then started to decrease from week 4, while for the somatic embryos derived from stem nodal section explants it increased up to week 10. This is in contrast with the heart stage somatic embryos (Figure. 8), which continued to increase from week 2 to week 10 in each explant derived from either leaf or stem nodal sections.
The fact that the number of globular stage somatic embryos from leaf explants started to decrease in week 4 of culture was probably due to the occurrence of the phase transition from the globular stage to the heart stage without increasing the number of globular stage somatic embryos. The number of globular embryos from the stem nodal explants continued to increase, presumably because the somatic cells in the stem nodal section explants were more induced to become IEDCs (induced embryonic determined cells) with a higher potential to become globular. It seems that the somatic embryos from stem nodal explants still developed, although some globular stage somatic embryos changed into heart stage somatic embryos.
Figure 9 shows that the torpedo stage of the somatic embryos started to emerge in the leaf explant-derived somatic embryos after 6 weeks of culture and in the stem nodal-derived somatic embryos after 4 weeks of culture. This is probably due to the fact that the regeneration ability of explants derived from stem nodal sections is higher than that of explants derived from leaf. Based on this research, explants derived from stem nodal sections produce a higher number and faster development of somatic embryos than leaf explants, therefore stem nodal section explants are more suitable as a source of explants for somatic embryogenesis in potatoes (Solanum tuberosum L.).
4 Conclusion
From this experiment we found that the explants with the highest potential to produce somatic embryos of potatoes (Solanum tuberosum) cv. Granola are stem nodal section explants and the optimum combination of plant growth regulators (2,4-D and BAP) to induce somatic embryos is 1.0 μM 2,4-D and 0.5 μM BAP. The highest maturation rate of somatic embryos of potato occurred in MS medium supplemented with 5 μM BAP.
Acknowledgements
This research was supported by a grant-in-aid from the Indofood Riset Nugraha 2012 program funded by PT Indofood Sukses Makmur Tbk.