
1 Introduction
Chitinases (EC 3.2.1.14) are a group of enzymes capable of degrading chitin, a long-chain linear polymer of N–acetyl glucosamine (GlcNAc), into its derivates by hydrolyzing the β-1,4-glycosidic bonds. Chitin is assumed to be the second most plenteous polysaccharide on earth [1]. In general, the enzymes responsible for chitin degradation are divided into endochitinases and exochitinases [2]. Endochitinase catalyze chitin degradation into (GlcNAc)2 or other chitoligosaccharides (COS). Exochitinases degrade chitin into GlcNAc. Chitooligosaccharides have been shown to have anti-tumor effects [3], antimicrobial effects [4], anti-fungal effects [5], anti-inflammatory effects [6] as well as the ability to increase bone strength [7]. Moreover, chitin derivates such as GlcNAc and D-glucosamine (GlcN) have been applied as nutraceutical
Received March 26th, 2015, 1st Revision October 6th, 2nd Revision February 15th, 2016, 3rd Revision May 16th, 2016, Accepted for publication May 19th, 2016.
Copyright © 2016 Published by ITB Journal Publisher, ISSN: 2337-5760, DOI: 10.5614/j.math.fund.sci.2016.48.2.6
agents for inflammatory bowel and osteoarthritis diseases [8]. In recent years, the demand for chitinase enzymes with new or desirable properties has increased due to the industrial application of COS and GlcNAc [9].
Chitinases are present in numerous organisms and recognized as extracellular inducible enzymes. Chitinase-producing bacteria can be isolated from various natural sources such as soil and aquatic habitats. Our previous research on isolation of thermopile microbes producing chitinase from Prataan Hot Spring was successfully conducted; several isolates of chitinolytic bacteria were successfully isolated. Among the isolates, the D3 showed the highest chitinolytic activity [10]. However, the optimum conditions for D3 isolate to produce chitinases and the characteristics of their chitinases have not been reported. Therefore, further research on the isolate is necessary.
In this study, identification of the D3 isolate, production optimization and characterization of the enzyme were carried out. The research involved identification of the isolate through 16S rRNA gene analysis followed by confirmation through morphological and physiological analysis; determination of optimum substrate, pH and temperature for chitinase production; determination of thermal stability and catalytic properties of the harvested enzyme at various temperatures and pH; and finally determination of the molecular weight of the purified chitinase.
2 Materials and Methods
2.1 Bacterial Strains and Cultural Medium
Thermophilic bacterium D3 isolated from Prataan hot spring in Tuban, East Java, Indonesia was used in this research. This isolate was identified as Paenibacillus sp. The isolate was grown in thermus colloidal chitin medium containing 0.7% (w/v) (NH4)2SO4, 0.1% (w/v) K2HPO4, 0.1% NaCl, 0.01% (w/v) MgSO4·7H2O, 0.05% (w/v) yeast extract, 0.1% (w/v) bacto tryptone and 0.5% colloidal chitin [11]. The chitin was produced from shrimp shell and the colloidal chitin was made in accordance with the report of Hsu & Lockwood [12].
2.2 Bacterial Identification
The isolate was identified through 16S rRNA gene sequencing analysis. The DNA was extracted from the pure culture using Fungal/Bacterial DNA MiniPrep Kit (Zymo Research). A partial DNA sequence for the 16S rRNA gene of D3 isolate was amplified using a pair of primers obtained from Macrogen F(5'CACGGATCCAGACTTTGATYMTGGCTCAG3') and
R(5'GTGAAGCTTACGGYTAGCTTGTTACGACTT5'). Amplifications were performed in 50 μl reaction mixture containing 25 μl of GoTaq Green Master Mix 2X (Intron), 5 μl of 16S forward primer 10 μM/μl, 5 μl of 16S reverse primer 10 μM/μl, 2.5 μl of DMSO, and 12.5 μl of double-distilled water (ddH2O). The mixture was subjected to the following amplification conditions with initial denaturation at 94 ºC for 5 min, 30 cycles of denaturation at 94 ºC for 45 sec, annealing at 55 ºC for 1 min, and elongation at 72 ºC for 1.5 min, as well as final elongation at 72 ºC for 5 min. The PCR products were purified using a PCR Purification Kit (Roche). The purified PCR products were then cloned into pGEMT-Easy (Promega) and transformed to competent E. coli DH5α cells. The presence of transformed cells was confirmed by colony PCR and DNA plasmid extracted from the transformed cells using a Plasmid Isolation Kit (Roche). This inserted plasmid was then sent to Macrogen, Korea for 16S rDNA sequence analysis.
The BLAST algorithm from GenBank was used to analyze the homology of the partial 16S rRNA gene sequence of the isolate [13]. Phylogenetic relationships were estimated using BLAST pairwise alignment (BLAST Tree). Bacterial confirmation and characterization through morphological and physiological properties were conducted according to Bergey's Manual of Systematic Bacteriology [14].
2.3 Optimization Chitinase Production
10% (v/v) of Paenibacillus sp. D3 isolate was inoculated into fermentation medium with the same composition as the cultivation medium. The medium containing the isolate was then agitated at 180 rpm (Yin Der shaker incubator). The basic conditions for fermentation were 0.9% (w/v) of colloidal chitin concentration, pH 7.0 and a temperature of 48 ºC. Determination of the optimum fermentation conditions was conducted by varying one parameter and controlling the other parameters. The fermentation time was varied from 0 to 72 hours, the colloidal chitin was varied from 0.7 to 1.1% (w/v), the pH was varied from 6.0 to 8.0, and the temperature was varied from 48 to 76 ºC. Sub-samples of the culture (50 mL) at initial and final fermentation were concentrated and analyzed for a chitinase activity assay. After determining the optimum fermentation time, the optimum concentration of colloidal chitin as well as the optimum fermentation conditions, reproduction of chitinase in large scale (1 L) was conducted for enzyme thermal stability assay, enzyme purification and characterization.
2.4 Enzyme Purification and Characterization
The enzyme was harvested from a 24 hour culture. The cells were precipitated by centrifugation (Thermo) at 12,000 g for 20 min at 4 °C and the cell-free supernatant, hereinafter called crude enzyme. The crude enzyme was then concentrated 30 times by freeze drying. The enzyme characterization was performed by varying the pH of buffer from 5.0 to 9.0 and the incubation temperature from 48 to 62 ºC during the activity assay. The thermal stability assay was carried out by pre-incubation heating of 0.6 ml concentrated enzyme in boiling water (100 ºC) for 0, 2, 10, 18, and 26 minutes. After the heattreatment, the enzyme was tested for chitinase activity.
Purification of the enzyme was conducted through several stages of purification. First, the concentrated enzyme was precipitated by 10% saturation of ammonium sulphate overnight at 4 °C. The precipitates were then obtained by centrifugation (12,000 g for 20 minutes) and dialyzed toward 0.02 M phosphate buffer pH 7.0 at 4 °C for 24 hours. Dialyzed enzyme solution was loaded into Gel Permeation Chromatography (GPC) Shepadex G-100 column (1 x 20 cm) (Sigma) equilibrated with 0.02 M phosphate buffer pH 7.0. The fraction showed the highest specific activity and the yield was prepared for further purification using IEC. The selected fraction was then loaded into a DEAE cellulose column (1 x 20 cm) (Sigma) equilibrated with 0.02 M phosphate buffer pH 7.0. The enzyme was eluted with linear gradient of NaCl (NaCl 0.1-1.0 M in 0.02 M phosphate buffer pH 7.0) at a flow rate of 0.5 mL/min. The eluted fractions were assayed for enzyme activity.
2.5 Chitinase Assay
The chitinase activity was measured by mixing 0.4 mL of concentrated or purified enzyme with 0.4 mL of colloidal chitin (0.1 g/ml), which was dissolved in 0.02 M phosphate buffer at pH 7.0. The mixture then was incubated at 48 ºC for 60 minutes [15]. The reaction was stopped by centrifugation at 12,000 g for 20 minutes. 0.6 mL of the supernatant was tested colorimetrically using the Nelson-Somogyi assay [16] by detecting the amount of GlcNAc present in the solution as a result of chitin degradation. N-acetyl glucosamine (Sigma) was used as the standard. The amount of enzyme which produces 1 mol of reducing sugar as N-acetyl-D-glucosamine equivalent per minute is defined as one unit of the chitinase activity (U). The Bradford colorimetric dye binding assay [17] was used for protein content determination and bovine serum albumin (BSA) was used as standard protein. The molecular mass of the purified chitinase was estimated by SDS-PAGE using a 10% gel, according to Laemmli [18]. The protein amount in each lane was controlled at approximately 20-30 µg. Protein bands were located by staining the gel with Coomassie Brilliant Blue R-250.
Chromatein prestained protein ladders in the range of 10-175 kDa (Vivantis) were used as standard protein molecular weight markers.
3 Results and Discussions
3.1 Identification and Characterization of Bacterial Isolate
Chitinolytic bacterium D3 was isolated from Prataan hot spring in Tuban, East Java, Indonesia. The homology of the partial 16S rDNA analysis confirmed that the isolate belonged to Paenibacillus sp. Phylogenetic relationships analysis using BLAST pairwise alignment showed that the isolate had maximum homology with Paenibacillus sp 9-7AIA (99% identity) (Figure 1). Paenibacillus species have been reported to have high activity of cell wall degrading enzymes [19] and chitinase [20], making this species commonly applied as bacteria and fungi biocontrol agents [21].
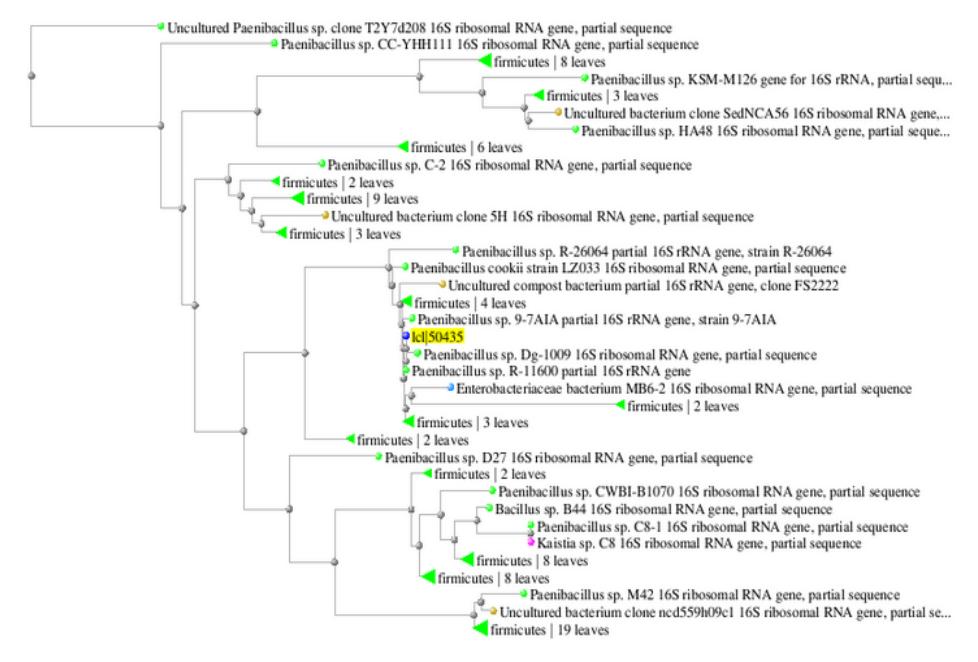
Figure 1 Phylogenetic tree based on 16S rDNA sequence analysis of D3 isolate showing homology with Paenibacillus sp.
The characterization assay through morphological and physiological analysis as presented in Table 1 corresponded to those presented in Bergey's Manual of Systematic Bacteriology [14]. The Paenibacillus sp. D3 showed a positive result in the following tests: catalase and oxidase production; acid production from glucose, xylose, arabinose and glycerol; the hydrolysis of starch and urea;
growth in 2-5% (w/v) NaCl; growth at 50 ºC; methyl red test; nitrate reduction, motility and anaerobic growth. The Paenibacillus sp. D3 showed a negative result in the following tests: acid production from manitol, hydrolysis of gelatin, utilization of acetate and citrate; growth in 7% (w/v) NaCl; indole formation; Voges-Proskauer test and nitrogen fixation.
Table 1 Morphological and physiological characteristic of Paenibacillus sp. D3.
| Characteristic | Colony Properties | ||
|---|---|---|---|
| Colony shape | Irregular | ||
| Elevation | Raised | ||
| Pattern-forming colonial growth | T (tip-splitting) | ||
| Colony color | Creamish white | ||
| Cellular morphology | Rod-shaped | ||
| Gram staining | Gram negative | ||
| Spore | Oval endospore | ||
| Catalase | + | ||
| Oxidase | + | ||
| Acid from: | |||
| - Glucose | + | ||
| - Xylose | + | ||
| - Arabinose | + | ||
| - Glycerol | + | ||
| - Mannitol | - | ||
| Hydrolysis of: | |||
| - Starch | + | ||
| - Gelatin | - | ||
| - Urea | + | ||
| Utilization of: | |||
| - Acetate | - | ||
| - Citrate | - | ||
| Growth in salinity | |||
| - 2 % NaCl | + | ||
| - 5% NaCl | + | ||
| - 7% NaCl | - | ||
| Indole formation | - | ||
| Methyl red test | + | ||
| Voges-Proskauer test | - | ||
| Nitrogen fixation | - | ||
| Nitrate reduction | + | ||
| Growth at 50 °C | + | ||
| Motility | + | ||
| Anaerobic growth | + | ||
The growth curve of Paenibacillus sp. D3 on thermus colloidal chitin medium (Figure 2) showed the lag, log, stationary and death phase observed during 0-4, 4-16, 16-32 and 36-48 hours incubation time respectively.
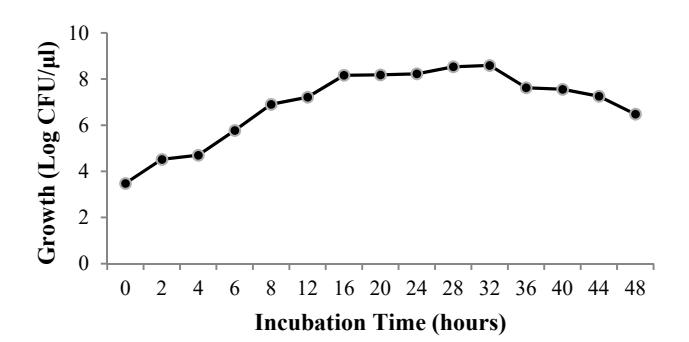
Figure 2 Growth of Paenibacillus sp. D3 (Log CFU/μl) in thermus colloidal chitin medium pH 7.0 for 48 hours at 48 ºC.
3.2 Optimization Chitinase Production
The incubation time required to reach the maximum enzyme level was determined by the characteristics of the culture as well as the growth and enzyme production rate. The effects of incubation time on chitinase production are shown in Figure 3(a). The Paenibacillus sp. D3 produced the highest chitinase at 24 h of incubation. The enzyme production decreased at 36 h and there was no significant difference (p-value > 0.05) in enzyme production from 36 h to 72 h during incubation. The decreasing production of the enzymes may be caused by the lack of nutrients or toxic substances produced, resulting in the inactivation of the enzymes [22]. With respect to cell growth, the highest chitinase production at 24 hours was in the middle of stationary phase. In this phase, accumulation of chitinase that had been produced since the exponential phase occurred. Compared to other microorganisms, Paenibacillus sp. D3 can produce chitinase optimally in shorter incubation times. As reported by Nawani [23] with Microbispora sp., Wang and Hwang [24] with B. cereus, B. alvei and B. sphaericus, as well as Saima, et al. [22] with Aeromonas hydrophila and Aeromonas punctata produced the highest chitinase yield after 48 h of incubation. Natsir, et al. [25] also reported that Bacillus sp. HSA,3-1a produced the highest chitinase yield at the end of the stationary phase at 72 h of incubation time.
Incubation temperature affects many biological processes, including the growth rate and enzyme production. To investigate the optimum temperature for chitinase production, a culture of Paenibacillus sp. D3 was grown at 48-76 ºC. Chitinase production reached maximum level at 48 ºC and the trend showed a decline from 55 to 76 ºC (Figure 3(b)). There was only a small amount of chitinase produced at 76 ºC. A different result was obtained when a different isolate was used. As reported by Natsir, et al. [25], Bacillus sp. HSA,3-1a produced the highest chitinase yield between 55 and 60 °C incubation temperature.
To investigate the effect of pH of media on the chitinase production, bacterial cultures were grown at different pH (6.0-8.0) (Figure 3(c)). Among the different pH of the media studied, pH 7.0 supported the maximum chitinase production of Paenibacillus sp. D3. However, there was no significant difference (p-value > 0.05) in chitinase production between pH 6.0 and 7.0. Similar observations were also reported by Karunya [26] with Bacillus subtilis, by Sowmya, et al. [27] with Streptomyces sp., by Natsir, et al. [25] with Bacillus sp., which produced the highest chitinase at neutral pH.
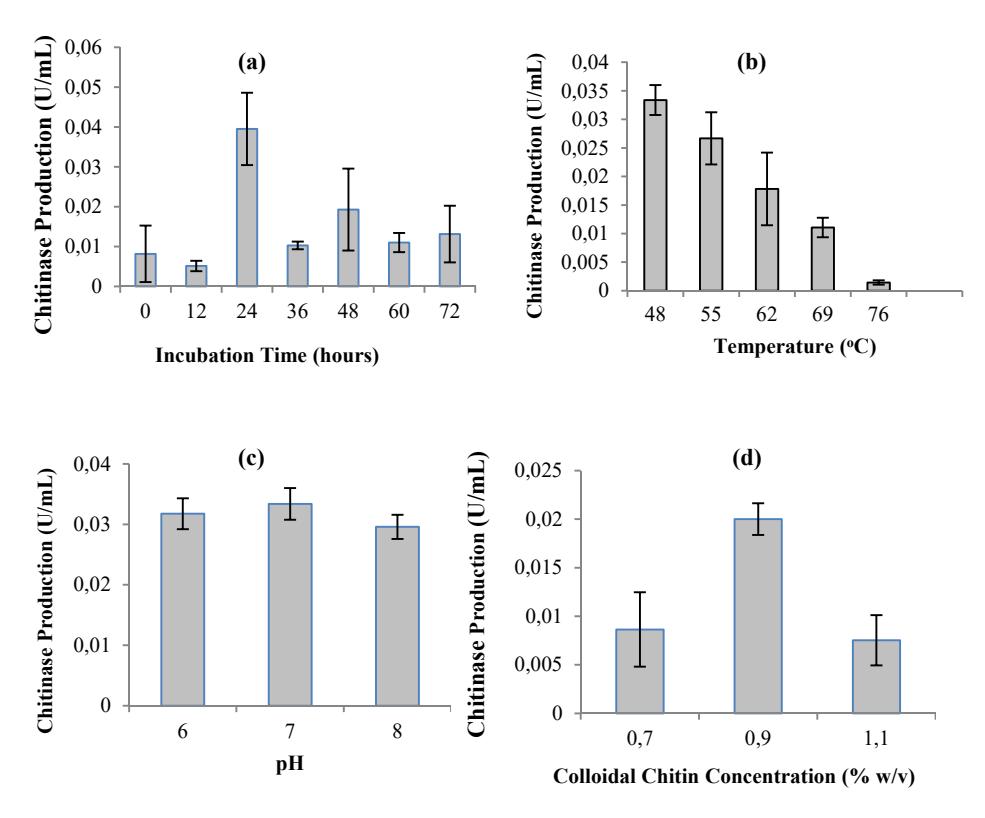
Figure 3 Effect of incubation period (a), temperature (b), pH (c) and colloidal chitin concentration (d) on chitinase production by Paenibacillus sp. D3. The basic conditions for fermentation were 0.9 % (w/v) colloidal chitin concentration, pH 7.0 and incubation at 48 ºC. Determination of the optimum fermentation conditions were conducted by controlling fermentation factors.
Another important factor that can induce chitinase production in several microorganisms is colloidal chitin concentration [28,22]. Experiments with various substrate concentrations were conducted to determine the optimum substrate concentration that can react with the enzyme. Figure 3(d) shows that the maximum chitinase production by Paenibacillus sp. D3 was achieved at 0.9% (w/v) of colloidal chitin concentration. The majority of the bacteria produced the maximum level of chitinase at 0.3% of colloidal chitin concentration as reported by Natsir, et al. [25], Karunya, et al. [26] and Saima, et al. [22]. Paenibacillus sp. D3 has slight similarity with Serratia marcescens and T. harzianum, which have been reported to have maximum chitinase production at 1% of colloidal chitin concentration [29].
3.3 Enzyme Purification and Characterization
Enzymes can function as acid, base or neutral based on their amino acid constitution in their active sites, which can be influenced by their environmental pH. The effect of pH on the activity of chitinase produced by Paenibacillus sp. D3 was evaluated. The chitinase showed a broad range of pH tolerance from 5.0 to 9.0 with optimum pH between 7.0 and 8.0, as shown in Figure 4(a). Regarding the stability of enzyme activity against different pH values, the chitinase retained more than 76% activity from pH 5.0 to 8.0 and retained 47% activity at pH 9.0. The aforementioned result on optimum pH is similar to that from Micrococcus sp. AG84 [30], Bacillus sp. LJ-25 [31] and Bacillus sp. HAS, 3-1a [25]. However, a different optimum pH was obtained by the chitinase enzyme isolated from Aeromonas sp. DYU-Too7 [32], Vibrio sp. 98CJ11027 [33] and Paenibacillus sp. D1 [34], which were mostly active at acidic pH. The optimum pH determined in this experiment was used for further experiments.
The effect of temperature on the enzyme activity was determined by measuring chitinase activity between 48 and 62 ºC. The optimum temperature of chitinase activity produced by Paenibacillus sp. D3 was 55 ºC, as shown in Figure 4(b). The chitinases produced from the isolate can be categorized into thermophilic and thermostable enzymes. A stability test conducted at 100 °C in the preincubation step showed that the enzyme still retained its activity by 51% after being heated for 26 minutes (Figure 4(c)). The result on optimum activity temperature was similar to that from Vibrio alginolyticus-H-8 [35]. However, a different optimum activity temperature was given by chitinase isolated from Paenibacillus sp. D1 and Bacillus sp. HSA,3-1a, which had optimum activity at 50 °C and 60 °C respectively [34,25].
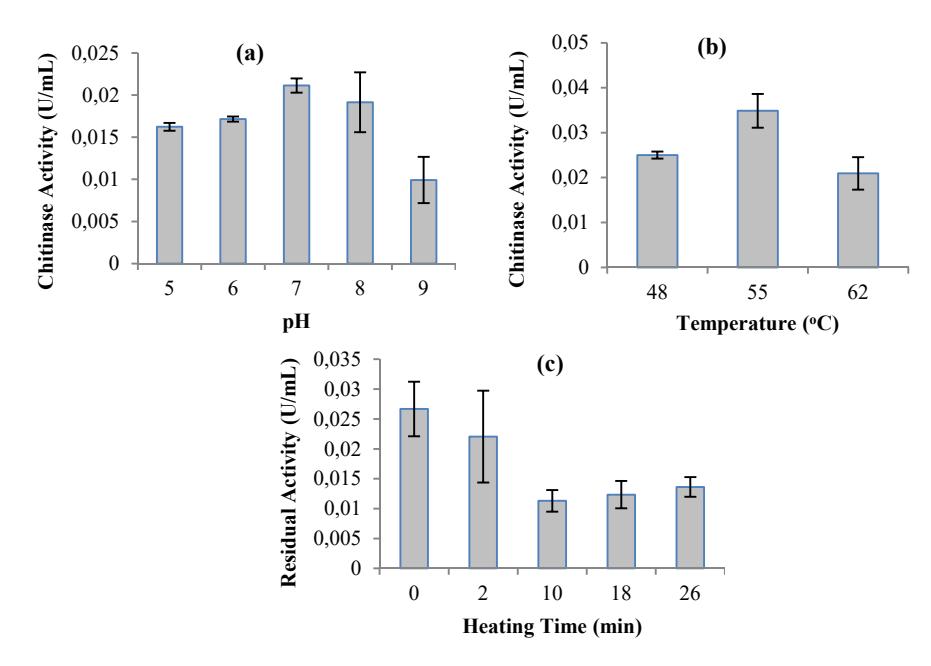
Figure 4 Characterization of chitinase produced by Paenibacillus sp. D3. (a) The effect of pH on the activity of chitinase. (b) The effect of temperature on the activity of chitinase. (c) Thermal stability of chitinase after heating at 100 ºC.
Table 2 Purification of Chitinase from Paenibacillus sp. D3 isolate.
| Purification step | Total Protein (μg) | Total Activity (U) | Specific Activity (U/μg) | Yield (%) | Purification (fold) |
|---|---|---|---|---|---|
| Concentrated enzyme | 8927.39 | 0.0437 | 4.89 x 10-6 | 100 | 1 |
| (NH4)2SO4 | 2263.71 | 0.0137 | 6.04 x 10-6 | 31.32 | 1.24 |
| Shepadex G-100 | 88.98 | 0.0038 | 4.26 x 10-5 | 8.68 | 8.71 |
| DEAE cellulose | 25.38 | 0.0012 | 4.61 x 10-5 | 2.68 | 9.43 |
Table 2 summarizes the results of each purification step of chitinase from Paenibacillus sp. D3. The chitinases were isolated from the concentrated culture supernatant by adding ammonium sulfate with 10% saturation. The results showed that the enzyme was purified 1.24-fold with a specific activity of 6.04 x 10-6U/µg protein after ammonium sulfate fractionation. The sediment of proteins was then dialyzed toward 0.02 M phosphate buffer pH 7.0, followed by gel filtration through Sephadex G-100. The GFC yielded three peaks, as shown in Figure 5. The highest chitinase activity was achieved by fraction 21, which was then used in further purification steps. After the GFC step, the enzyme was purified 8.71-fold with specific activity of 4.26 x 10-5 U/μg protein. Fraction 21 from the GFC step was then purified by IEC using DEAE cellulose.
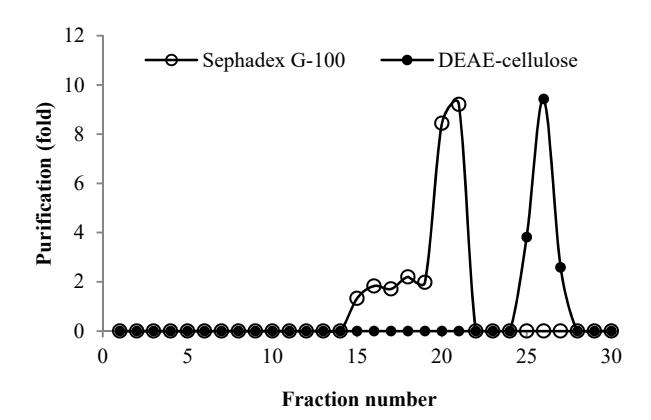
Figure 5 Chromatogram of chitinase from Paenibacillus sp. D3 on Sephadex G-100 (o) and DEAE cellulose (●).
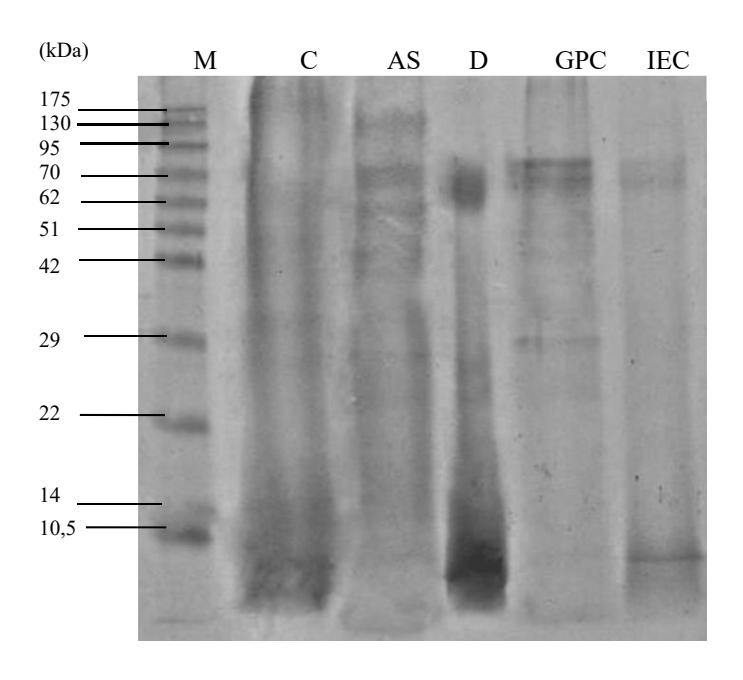
Figure 6 SDS PAGE analysis of chitinase at each purification step. M: protein marker. C: concentrated enzyme. AS: ammonium sulfate precipitation. D: dialysis. GPC: Gel Permeation Chromatography using Sephadex G-100. IEC: ion exchange chromatography using DEAE cellulose.
The IEC step yielded one peak with highest chitinase activity at fraction 26 (Figure 5). The final purification step exhibited 9.43-fold enzyme purification with a specific activity of 4.61 x 10-5 U/µg protein. The results showed that the effectiveness of the purification method applied in this experiment was 9.43 fold with a 2.68% yield (Table 2). One of the methods that is considered to be very efficient in separation and resolution of protein components in the sample is SDS-PAGE. The results of the SDS-PAGE analysis at each purification step can be seen in Figure 6. Two protein fractions were obtained from SDS-PAGE with molecular weights of 68 and 82 kDa. These results are similar to previously reported data from another Paenibacillus sp., where the chitinases had molecular masses ranging from 40 to 150 kDa [34,36].
4 Conclusion
The D3 isolated in this research belonged to Paenibacillus sp., which is a novel thermophilic bacterial strain that produces the highest level of chitinase at 0.9% (w/v) of colloidal chitin (pH 7.0) at 48 °C for 24 h. The chitinase has two protein fractions with molecular weights of 68 and 82 kDa. With a broad range of pH and thermal stability, the chitinase has potential applications in recycling chitinous wastes.
Acknowledgements
This work was supported by the Research Fund from the University of Surabaya with contract number 087/Lit/LPPM-01/FTB/VIII/2013.