
1 Introduction
Breast cancer is the leading cause of cancer deaths among women worldwide. The GLOBOCAN project of the International Agency for Research on Cancer (IARC) states that breast cancer incidence is highest in Indonesia. In 2008 there were approximately 156,500 new cases of breast cancer and 104,500 deaths due to breast cancer [1]. Cancer development involves many processes, including angiogenesis and metastasis. Angiogenesis is regulated by angiogenic stimulators such as HIF-1α protein, while focal adhesion kinase (FAK) and Wnt2 protein play an important role in metastasis. It is known that HIF1α contributes to the process of angiogenesis and is responsible for cell survival under conditions of hypoxia [2]. Wnt2 contributes to the EMT (epithelial mesenchymal transition) process and accumulation of beta-catenin in the nucleus, which acts as a transcription factor targeting genes that play a role in the metastatic process, and FAK plays a role in the formation of focal adhesions and interacts with integrin [3,4]
Nowadays many cancer therapies are focused on blocking signaling pathways that are important in the development of cancer. In this research we observed natural products that have potential to be used for combating cancer. They presumably block several proteins in signaling pathways involved in angiogenesis and metastasis. Propolis has antimicrobial, anti-inflammatory, and antitumor activity. Propolis is able to suppress cancer-induced angiogenesis through VEGF [5] and to inhibit migration of human umbilical vein endothelial cells (HUVEC) [6]. Moreover, it is able to induce apoptosis in breast cancer cell line MCF7 at a concentration of 47.45 mg/ml [7]. α-mangostin is a derivative of xanthone contained in the mangosteen fruit pericarp. Mangostin has several biological activities, such as inhibition of HIV-1 protease, antibacterial activity and stimulation of phagocytic cells, and can induce apoptosis in cancer cells HL60 [8]. Both of these natural products have been widely tested in vitro, but more testing is still needed to obtain a description that can represent the effect of both natural products separately or of a combination of both products in in vivo conditions.
The diversity of pathways that can be activated or inhibited in the process of cancer development often leads to ineffectiveness of cancer therapies. Therefore, it is important to combine natural products that have different effects on cancer development. Until now, the effects of natural products such as propolis and mangostin against cancer development have not been observed. The aim of this study was to observe the effect of propolis and mangostin on mammary gland cancer development and on the expression of Wnt2 and FAK in Balb/C mice.
2 Materials and Methods
2.1 Tumor Induction in Mice and Treatment by Combination of α-Mangostin and Propolis Extracts
Mice (Balb/C), purchased from LPPT Gadjah Mada University, Yogyakarta, were maintained in a 12-hour light dark cycle. The mice were provided with food and water ad libitum and were used at 9 weeks of age. DMBA was induced 11 times in the mice after oral administration for 5 weeks. Treatment with propolis and α-mangostin was performed after the tumor reached a size of 200 mm3 [9]. The treatment of the mice was approved by the Research Ethics Committee, Faculty of Medicine, Maranatha Christian University (Decree No. 170/KEP/V/2014).
In this study, 25 female mice Balb/C were divided into 5 groups. The mice in group K- served as negative control. The mice in group M were treated with αmangostin (M-002, Indofine Chemical Company, Inc.) with a concentration of 0.128 mg/kg body weight. The mice in group P were treated with propolis (RAHMI apitherapy) with a concentration of 0.32 mg/kg body weight. The mice in group MP were treated with mangostin (0.128 mg/kg bw) combined with propolis (0.32 mg/kg bw). The positive control group (Dx) was treated with doxorubicin (anti-cancer drug) at 14.04 mg/kg body weight. The αmangostin dose given to the mice was determined based on IC50 of mangostin in the MCF-7 cell line from our previous study [7], namely 1.6 mg/mL. The propolis dose given to the mice was determined based on IC50 of propolis in the KATO III cell line, i.e. a concentration of 4.09 mg/mL [10].
Alpha-mangostin and propolis were administered orally every day for 14 days. Doxorubicin was given intravenously to the Dx group once every 2 days over a span of 14 days. On the 15th day after treatment, the mice were necropsied using ether in a sealed container. The mice were then dissected and the tumors were collected and put into a container. The tumors were washed and cleaned with cold PBS three times and stored in a cryotube, sealed, labeled, and stored in liquid nitrogen.
2.2 Preparation of Whole-cell Lysates and Nuclear Extracts
Hundred (100) mg of tumor tissue was lysed with 400 μl buffer lysis solution A (10 mM HEPES pH 7.9, 10 mM KCl, 0.1 mM EDTA pH 8, 0.1 mM EGTA pH 8) and then incubated on ice for 10 minutes. Furthermore, NP-40 was added to the lysate, which was further vortexed for 10 seconds. The suspension was centrifuged for 1 minute at 13,000 rpm at 4 °C and then supernatant I (containing cytoplasmic protein) was collected.
The nuclear pellet was re-suspended in nuclear extract buffer (20 mM HEPES pH 7.9, 400 mM NaCl, 1 mM EDTA pH 8, 1 mM EGTA pH 8, 25% glycerine). The nuclear suspension was incubated on ice for 15 minutes and then centrifuged at 13,000 rpm for 5 minutes. Supernatant II (contained soluble nuclear fraction) was stored at -80 °C. The protein concentration was determined with the Bradford protein assay.
2.3 Western Blotting Analysis
The proteins (50 µg protein) were electrophoresized on 10% SDS-PAGE gels and then transferred onto PVDF membranes using a Mini Trans-Blot. Nonspecific binding on the membranes was blocked with 5% (w/v) nonfat skim milk for 1 hour. The membranes were washed with TBST (Tris-Buffered Saline and Tween-20) five times for 10 minutes and incubated with an appropriate
dilution of specific primary antibodies in TBST overnight at 4 ºC. Anti-mouse HIF1α (Abcam ab463, 1:2000), mouse anti-FAK (Abcam ab40794, 1:6000), anti-mouse Wnt2 (Abcam ab109222, 1: 2500), and mouse Anti-Actin (Abcam ab1801, 1:1000) were used as primary antibodies. Subsequently, the membranes were washed with TBST and incubated with the appropriate secondary antibody (anti-mouse IgG-POD, Fab fragments/anti-rabbit IgG-POD (Roche 11520709001, 1:5000) for 1 hour. The membranes were further washed five times for 10 minutes in a TBST. Specific protein detection was conducted by enhanced chemiluminescence.
3 Results and Discussion
3.1 Effect of Propolis and Mangostin on Mice Balb/C Body Weight
The body weight of the mice in the control group (K-) increased throughout the 2-week observation period, while that of the positive control mice (Dx), the mangostin group mice (M), the propolis group mice (P) continued to decrease during this period, as shown in Figure 1. Interestingly, the body weight of the MP (mangostin and propolis) group mice did not change much during the period of observation. This suggests that administration of propolis together with mangostin has no effect on the body weight of Balb/C mice. This indicates that propolis and mangostin are safe for consumption and do not cause side effects.
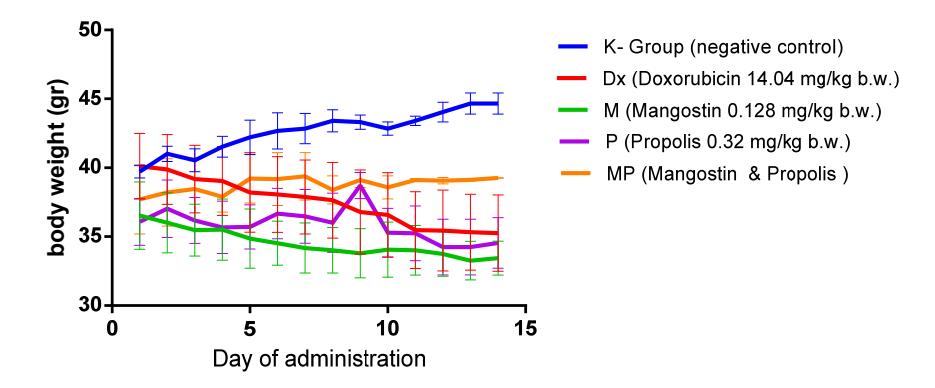
Figure 1 Effect of mangostin and propolis on Balb/C mice body weight.
Similar results have also been reported by Sugimoto, et al. [11], i.e. the administration of propolis did not affect the body weight of induced lung cancer
mice. These results show that the bioactive compound, artepillin C, in propolis has no detrimental effect.
Our result showed that mangostin is not toxic and can be used as an alternative cancer treatment. Kosem, et al. [12] showed that Balb/C mice implanted with colon cancer in the NL17 cell line and treated with mangostin at a dosage of 200 mg/kg did not lose weight during treatment.
3.2 Effect of Propolis and Mangostin on Mammary Cancer Development in Balb/C Mice
Our results show that administration of mangostin (0.128 mg/kg bw) as a single compound is unable to inhibit cancer development in vivo. However, extracts of propolis (0.32 mg/kg bw) were able to decelerate cancer progression in Balb/C mice, mostly when propolis was administered together with mangostin (MP group). The development of cancer in the MP group was retarded more than in the Balb/C mice treated only with propolis, as shown in Figure 2. This tendency shows that there are probably some interactions between both natural ingredients that inhibit tumor progression.
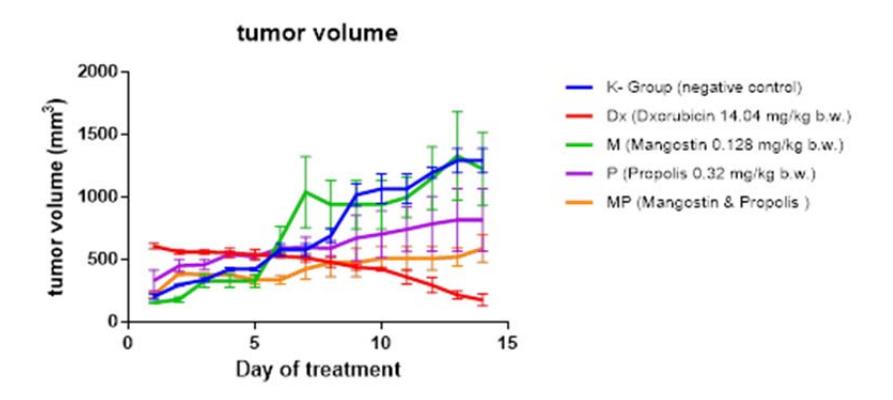
Figure 2 Effect of mangostin and propolis on Balb/C mice tumor development.
The low effect of single-compound treatment with mangostin or propolis on tumor development in Balb/C mice is assumed to be affected by the low dose of the mangostin (0.128 mg/kg bw) as well as propolis (0.32 mg/kg bw). Mouse, et al. (2012) have shown that the administration of propolis at a dosage of 2.5 mg per mouse can reduced tumor volume significantly. Additionally, Shibata, et al. [13] have shown a significant inhibition of tumor growth in female Balb/C mice that were implanted with BJMC3879luc2 cells and treated with mangostin 20 mg/kg bw.
The inhibitory effect of mangostin combined with propolis on tumor progression is allegedly caused by the ability of mangostin and propolis to reduce free radicals. Free radicals, which are produced abundantly in cancer tissue, can be reduced by mangostin and propolis. Sun, et al. [14] and Padmavathi, et al. [15] have shown that mangostin and propolis are both antioxidants.
3.3 Pattern of HIF-1α Protein Expression, FAK, and WNT2 on Mammary Tumors in Balb/C Mice
In this study, expression patterns of genes that play a role in the process of angiogenesis (i.e. HIF-1α) and metastasis (Wnt2 and FAK) were observed (Figure 3). Our results show that administration of mangostin combined with propolis inhibits Wnt2 expression in mammary tumors. This inhibition is also expected to have an impact on the tumor metastasis mediated by Wnt2 proteins.
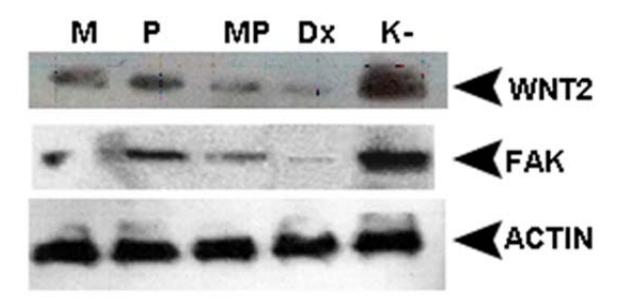
Figure 3 Western blot results of Wnt2 and FAK in tumor tissue of mice treated with mangostin and propolis. K- is the negative control group. Dx is the positive control group, which was given doxorubicin 14.04 mg/kg bw P is tumor tissue from mice that were treated with propolis (0.32 mg/kg bw). M is tumor tissue from mice that were treated with mangostin (0.128 mg/kg bw). MP is tumor tissue from mice that were treated with propolis (0.32 mg/kg bw) and mangostin (0.128 mg /kg bw).
These results are also supported by Mahmoud, et al. [16], who state that caffeic acid phenethyl ester (CAPE), one of the active compounds in propolis, inhibits the growth of colorectal cancer cells by decreasing the expression of betacatenin in the nucleus and increases the expression of p53, which leads to apoptosis. β-catenin is included in the downstream canonical pathway of Wnt2 [17]. Additionally, Chitchumroonchokchai, et al. [18] have shown that the tumor mass and concentration of Bcl-2 and β-catenin in tumors of mice fed with an α-mangostin diet were lower than in mice fed with the control diet.
Similar with Wnt2, FAK was downregulated in the MP group and the expression of FAK in MP group was lower than in groups M and P and was closely similar to the Dx group (Figure 3). This inhibition is also expected to have an impact on the mechanisms of tumor metastasis mediated by the FAK protein. This result is supported by Shih, et al. [19], who have shown that αmangostin can inhibit αvβ3 integrin, focal adhesion kinase (FAK), ERK1/2, MMP-2 and MMP-9.
HIF1α expression in group MP as well as in group Dx was totally inhibited, while in group M, HIF1α was downregulated. However, in group P the expression of the HIF1α protein was greater than in groups K- and M (Figure 4). It is suspected that the active compounds contained in propolis have a low inhibitory effect on HIF1α expression. Hattori, et al. [20] have reported a similar result, which indicates that some of the active ingredients in propolis, isosakuranetin and beturetol can induce the expression of HIF-1α target genes and HIF1α activity in hypoxic conditions.
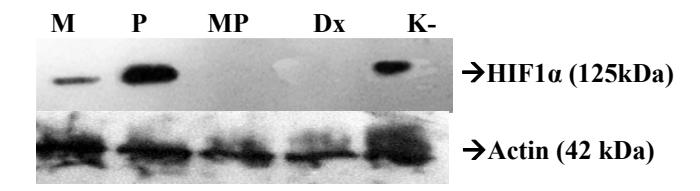
Figure 4 Western blot results of HIF-1α in tumor tissue of mice treated with mangostin and propolis. K- is the negative control group. Dx is the positive control group, which was given doxorubicin 14.04 mg/kg bw P is tumor tissue from mice that were treated with propolis (0.32 mg/kg bw). M is tumor tissue from mice that were treated with mangostin (0.128 mg/kg bw). MP is tumor tissue from mice that were treated with propolis (0.32 mg/kg bw) and mangostin (0.128 mg /kg bw).
The results show that the combination of propolis and mangostin suppressed the expression of Wnt2, FAK and HIF1α. Alpha-mangostin and active compounds contained in propolis supposedly interact with proteins that act on a target protein, such as p53. Paulino, et al. [21] stated that the active compounds contained in propolis, such as Artepillin C or CAPE, can increase the expression of p53, which plays a role in the apoptosis process. Watanapokasin, et al. [22] have shown that expression of p53 protein increased in the COLO205 cell line after treatment with 20 μg/ml of mangostin for 3 hours. Chinese propolis extract caused an increase of p53 mRNA levels in HCT116 cell lines [23]. Therefore, both propolis and mangostin may cause p53 to be retained in the cell and to be bound to Mdm2 leading to HIF1α degradation. Kaur, et al. [24] mention that p53 modulates Mdm2 by adding ubiquitin to HIF1α proteins, which affect HIF1α degradation. On the other hand, retention of p53 in cancer cells may have suppressed FAK in the MP group in this research. Golubovskaya, et al. [25] have shown that FAK regulates p53 protein in cancer cells.
The inhibition mechanism of propolis and mangostin on Wnt expression is still unclear, but it is assumed that mangostin and propolis can suppress the expression of Wnt in Balb/C mice, which subsequently leads to the decline of βcatenin as a result of β-catenin degradation. The degradation of β-catenin can be due to increased expression of p53 in cancer cells of the mice in the MP group. Sadot, et al. [26] state that p53 wild-type can decrease β-catenin expression in human and mice cells.
4 Conclusions
Administration of a combination of propolis and mangotin as an alternative treatment as part of cancer therapy is a safe therapy in view of the insignificant effect of both treatments on the body weight of the Balb/C mice. The combination of mangostin and propolis is able to inhibit cancer development and has potential anti-angiogenic and anti-metastatic activity due to a downregulated expression of Wnt2, FAK, and HIF1α in the cancer tissue.
Acknowledgements
This study was funded by Program Riset dan Inovasi ITB 2013, No. SITH. PN-6-09-2013 and BOPTN.