
1 Introduction
Foxtail Millet (Setaria italica (L.) Beauv.) belongs to the family of Poaceae. This millet is native to China. In East Asia and Southeast Asia, foxtail millet was used as a staple food prior to the introduction of rice. Foxtail millet has many benefits. The seeds have been used as raw material for beer in Europe, while in India, China, Japan, and Africa, foxtail millet seeds are used as ingredient for foods such as pudding, bread, cake, noodles and porridge [1]. In Indonesia, however, the use of foxtail millet as a food source is still limited. The usage of foxtail millet seeds as a rice substitution is limited to Buru Island [2]. The nutrient content of foxtail millet seeds is higher than that of rice. 100 g of dried foxtail millet seeds contain 351 kcal, 6.7 g crude fiber, 11.2 g protein, 4.0 g fat, 63.2 g carbohydrates, 31 mg calcium (Ca), 2.8 mg iron (Fe), 0.59 mg B1 vitamin (thiamine), 0.11 mg B2 vitamin (riboflavin) and 3.2 mg niacin [3].
Attempts to regenerate foxtail millet via in vitro culture have been conducted by several researchers [4-6]. Regeneration of foxtail millet has been done through shoot regeneration as well as somatic embryogenesis [4]. However, in vitro regeneration of foxtail millet cultivar Buru hotong has never been reported. Therefore, this research proposes the first protocol for in vitro regeneration of foxtail millet cultivar Buru hotong to facilitate future foxtaill millet breeding. Rout and co-workers [4] have reported that in vitro regeneration of foxtail millet can be obtained using a mesocotyl as an explant grown on MS media supplemented with 6-benzylaminopurine (BAP), kinetin, 2,4 dichlorophenoxyacetic acid (2,4-D) and 1.5 ppm NiSO4. In this paper, we further improve the in vitro regeneration protocol for foxtail millet cultivar Buru hotong (Setaria italica) through direct and indirect organogenesis.
2 Materials and Methods
2.1 Shoot Induction and Multiplication
Leaf base of 10-day old seedlings was isolated and used as explant. This explant was derived from foxtail millet seeds germinated on MS basal media [7] without the addition of plant growth regulators (PGRs). For shoot induction, the explants were cultured in MS basal media, supplemented with various combinations of 2,4-D, kinetin, and BAP (Table 1), 1.5 ppm NiSO4, 3% sucrose and 2.5% gelrite (with pH 5.9). The leaves were incubated in a 12/12 h light/dark photoperiod at 25 ºC. The induced shoots were then transferred into MS basal media without the addition of PGRs (MS 0) for shoot multiplication.
| BAP (ppm) | ||||
|---|---|---|---|---|
| KIN (ppm) | 0.5 | 1.0 | 2.0 | 2,4-D (ppm) |
| 0.5 | A | B | C | 0.1 |
| 1.0 | D | E | F | 0.5 |
| 2.0 | G | H | I | 0.6 |
Table 1 Combinations of PGRs used in media for shoot induction.
2.2 Root Induction and Plantlets Acclimatization
Roots were induced by transferring shoots from shoot multiplication media into root induction media consisting of MS basal media without PGRs. The shoots were incubated in a 12/12 h light/dark photoperiod at 25 ºC. Prior to acclimatization, the plantlets were placed in a room with light intensity above 1000 lux for 7 days of hardening. The plantlets were then removed from the bottle and rinsed with tap water, followed by soaking in 1.5 g/L of dithane (fungicides) and 1 g/L of bactericide for 10 min. The plantlets were then planted into growing media consisting of rice husk charcoal, coco peat, or mixed media
(containing rice husk charcoal, humus and coarse fern powder in a ratio of 1:2:1).
3 Results and Discussion
The explants showed a response after one week of culture in shoot induction media. The responses were in the form of callus or direct organogenesis (shoots). PGR combination in media D, E, G, H, and I (Table 1) was able to induce callus formation. A high percentage of callus was formed in MS basal media containing 1 ppm kinetin + 1 ppm BAP + 0.5 ppm 2,4-D (media E), in which around 73% of explants formed callus (Table 2). This result is consistent with the results of Rout, et al. [4], where mesocotyl regenerated a high frequency of embryogenic calli in a medium containing 1 ppm kinetin, 1 ppm BAP and 0.5 ppm 2,4-D. Callus formation is influenced by the interaction between auxin and cytokinin. Auxin and cytokinin have a synergistic role in cell division. Auxin causes differentiated cells (explant) to become dedifferentiated cells (callus). Cell dedifferentiation was probably caused by reorganization of the chromatin structure, which altered gene expression patterns. Changes in this expression pattern lead to the formation of callus [8]. On the other hand, cytokinin plays a role in activating genes that are responsible for cell division [9].
A yellowish-white compact callus (Figure 1a) was detected in the media containing kinetin (1 and 2 ppm), 2,4-D (0.5 and 0.6 ppm) and BAP (0.5, 1, and 2 ppm) (Table 3). The calli in media G, H and I, however, immediately turned brown after several days in culture (Figure 1c). Gaba [10] states that at higher concentrations, auxin will stimulate ethylene formation, yet causes senescence. The growth of senescent callus is usually slow and it will finally turn brown. Tang and Newton [11] have reported that browning callus reduces its regeneration ability. Four weeks after culture, some greenish area was formed on the surface of the callus grown in media E (Figure 2b), followed by shoot formation after several weeks in culture. According to Armini, et al. [12], the green area on the callus surface indicates cell differentiation that will lead to shoot formation.
Table 2 Percentage of explants that formed calli in induction media.
| Period of culture | ||||
|---|---|---|---|---|
| Media | 1st week | 4th week | ||
| D | 57.14 + 13.80 | 65.71 + 19.02 | ||
| E | 71.67 + 18.01 | 73.33 + 15.57 | ||
| G | 61.43 + 14.60 | 61.43 + 14.60 | ||
| H | 58.00 + 27.41 | 66.00 + 26.33 | ||
| I | 66.67 + 19.69 | 66.67 + 19.69 | ||
Figure 1 (a) Compact calli, (b) Indirect shoot organogenesis developed on greenish area of callus, (c) Browning calli.
In addition to callus formation, direct shoot organogenesis was observed in this research (Figure 2). Shoots were induced directly when the explants were cultured in media A, B, C and F (Table 3). The different morphological responses of the explants was probably caused by the different PGRs used in the media.
Figure 2 Shoots were induced directly on organogenetic induction media.
Table 3 Morphological response of Foxtail Millet explants on shoot induction media.
| Media | Morphological response | |
|---|---|---|
| A | Shoot | |
| B | Shoot | |
| C | Shoot | |
| D | Callus | |
| E | Callus | |
| F | Shoot | |
| G | Callus | |
| H | Callus | |
| I | Callus | |
Gunawan [13] explains that different levels of PGRs will affect growth and morphogenesis of explants. Furthermore, Close and Gallagher-Ludeman [14] have reported that different concentration levels of PGRs influence the balance of endogenous hormones in the explant and as a consequence it will regulate a different gene expression. Based on research conducted by Skoog and Miller in 1957, a higher ratio between cytokinin and auxin will lead to bud formation, while callus formation occurs when the auxin and cytokinin concentrations are in balance [15].
The highest number of shoots formed on MS basal media containing 0.5 ppm kinetin + 2 ppm BAP + 0.1 ppm 2,4-D (media C) (Table 4). It was found that media C was the most suitable media for direct shoot formation. Alizadeh, et al. [16] showed that direct and optimum shoot formation also occurred in wheat during culture in MS basal media supplemented with 2 ppm BAP. It seems that BAP is a suitable plant growth regulator for shoot induction. As stated by Van Staden, et al. [9], BAP is a plant growth regulator effective for in vitro shoot proliferation in several plants. This is probably due to the benzyl groups in BAP, which cannot be easily destroyed by enzymes in plants. In this research, shoots were formed mainly in the media containing higher concentrations of cytokinin than auxin. A similar result was yielded in the experiment of Marin-Mendez, et al. [17], who showed that in vitro shoot formation of maize (Zea mays) occurred in media containing more cytokinin than auxin. Liu and Bao [18] discovered that higher concentrations of cytokinin than auxin are more effective for shoot formation in Platanus acerifolia.
Table 4 Average number of shoots formed directly per explants in organogenesis induction media, 1 week and 4 weeks after culture.
| Number of shoots | |||
|---|---|---|---|
| Media | 1st week | 4th week | |
| A | 4.60 + 0.89 | 23.40 + 2.28 | |
| B | 2.20 + 1.30 | 25.60 + 2.07 | |
| C | 11.20 + 1.30 | 30.40 + 1.52 | |
| F | 6.20 + 0.84 | 15.40 + 2.41 | |
According to Shen [19], cells in explant tissue can dedifferentiate and directly form new primordial shoots. These cells become competent and are then able to respond to specific signals for shoot induction, e.g. hormones. Our results showed that competent cells were induced in the explants grown on organogenetic induction media. These competent cells were characterized by their ability to conduct cell division and then form shoots directly. Competent cells will be induced when the explant is grown on media containing auxin and cytokinin. Auxin induces cell division, elongation and differentiation because this growth regulator alters the pattern of gene expression, whereas cytokinin is responsible for increasing the sensitivity of cells to form shoots. In addition, cytokinin is also important for cell division [20]. Su, et al. [15] clarify that cell division is important for the formation of the initial cell in primordial shoot development.
Figure 3 Shoot multiplication and elongation.
Shoot multiplication occurred when shoots from shoot induction media had transferred into MS basal media without PGRs. Numerous shoots as well as elongated shoots were developed in this medium (Figure 3). Numerous cases of shoot multiplication mainly developed in the single shoot that was derived from direct shoot organogenesis (Figure 4).
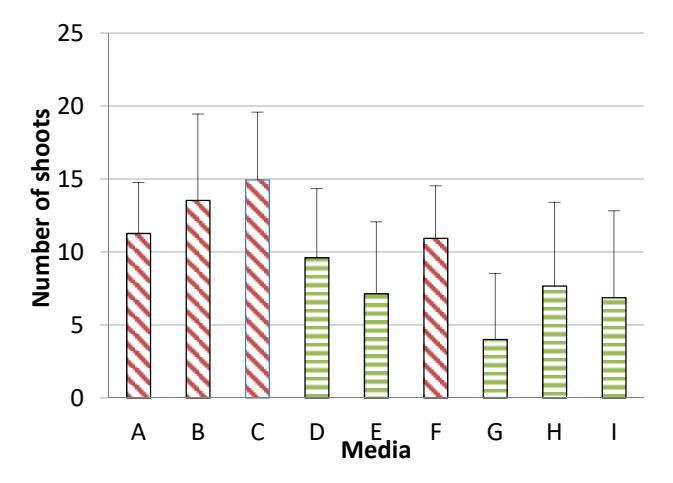
Figure 4 Shoots derived from callus (D, E, G, H, I) and shoots formed directly (A, B, C, F).
This is probably due to the presence of competent cells in shoot cells that were ready to form new primordia. Shoot multiplication also occurred in callusderived shoots, however, these shoots required a longer period to multiply because shoots produced in the callus phase requires competent cells to form and multiply shoots. Cell competency depends on hormones inside the callus tissue [21]. This result is in accordance with the result of Saeed, et al. [22]. He states that the multiplication of shoots derived from direct shoot organogenesis occurred in a shorter period than shoots formed in the callus phase.
Formation and elongation of new shoots were also influenced by auxin and cytokinin. In this research, the concentrations of endogenous auxin and cytokinin in the shoots seemed enough for shoot multiplication, because multiplication was done on MS basal media without PGRs. Cytokinin induces the formation of new shoots by stimulating genes that play a role in the formation of shoots and inhibits shoot apical dominance [23]. Cytokinin also plays a role in stimulating cell division by increasing the rate of protein synthesis, whereas auxin stimulates the elongation of the cells, thus causing bud and stem elongation.
Roots were also induced in this media (Figure 5). The highest percentage of root formation was obtained from direct organogenesis-derived shoots. Root initiation and formation require auxin. In the present experiment, however, roots developed from the basal shoots in the media without growth regulators. This was probably due to the in vitro shoots as well as the young leaves having produced auxin. Synthesized auxin will be transported through the cells of the leaves to the basal part of shoots [24].
Figure 5 Root formation in MS media without plant growth regulator.
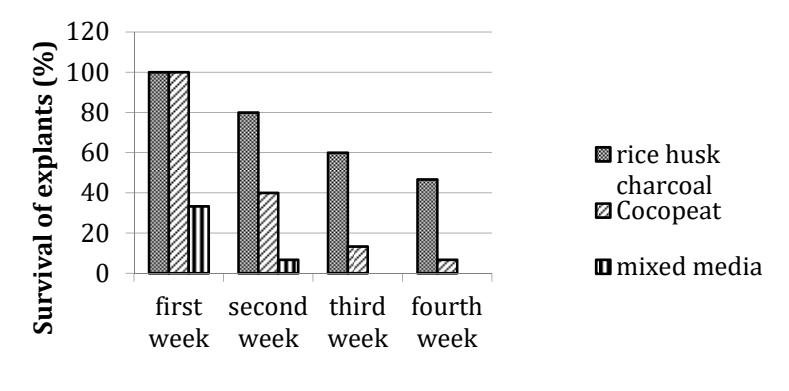
Acclimatization in three different growing media showed different survival rates of plantlets. Plantlets that were planted in rice husk charcoal media showed a higher survival rate (47%) compared to the other media. Plantlets were able to develop new leaves as well as new roots from stem nodes in the second week after planting (Figure 6). Rice husk charcoal plays a role in supporting the success of acclimatization by encouraging the growth and development of roots. Rice husk charcoal has good porosity, so roots can grow well. Good rooting allows the avaibility of nutrients and water for photosynthesis. In addition, rice husk charcoal can absorb and store water during the acclimatization and is also able to suppress growth of both fungal and bacterial pathogens [25].
Time after planting
Figure 6 Survival of plantlets on media rice husk charcoal, cocopeat and mixed media.
4 Conclusion
Based on the present results, it can be concluded that the optimum media for shoot stimulation in Setaria italica cv. Buru hotong was MS basal media containing 0.5 ppm kinetin, 2 ppm BAP, 0.1 ppm 2,4-D. Shoot multiplication and root induction were successfully developed in MS basal media without plant growth regulators. Acclimatization was better using rice husk charcoal media.