
1 Introduction
The Osteoarthritis (OA) is a chronic disease of the joints and bones due to trauma or other joint-related diseases [1]. OA is also associated with synovial inflammation, which causes disturbance of joint homeostasis [2]. Until now, an effective pharmacotherapy that can repair the function and the original structure of the damaged articular cartilage has not been found. Current therapies for OA use anti-inflammatory medication such as nonsteroidal anti-inflammatory drugs (NSAIDs). Biological therapies for OA and other orthopedic disorders are still being investigated [3]. Mesenchymal stem cells (MSCs) have been selected for OA therapy, because they are able to differentiate into chondrocytes and have the ability to repair articular cartilage. MSCs can be isolated from various types of mature tissue without reducing the potency of differentiation [3]. The aim of this research was to find an OA cell line model that can be used on treatment with MSCs in OA therapy.
In this research, human chondrocyte cells (CHON-002) were treated with IL-1β as a model of OA. IL-1β also promotes OA occurrence through several mechanisms: imbalance between damage and recovery of cartilage, reactive oxygen species (ROS), including nitrite oxide (NO), and inflammatory mediators such as prostaglandin E2 (PGE-2) through increasing of cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS) [4]. Free radicals induce oxidative stress, which can lead to an imbalance in both prooxidants and antioxidants, which further results in joint damage in patients with OA and rheumatoid arthritis (RA) [5].
Matrix metalloproteinases (MMPs) play a major role in the processes of chondrolysis. They are responsible for degenerative changes in the OA cartilage [6,7]. The messenger RNA (mRNA) identified in human cartilage, based on recent studies, comprises MMP-1, MMP-3, MMP-9, and MMP-13 [8,9], type II collagen degradation products by collagenase and also specific MMP proteins [10-12]. This type of enzyme is related to an intrinsic chondrocyte-mediated degenerative change of the cartilage matrix in OA. However, the importance of these enzymes remains unclear. The activities of metalloproteinase can be inhibited by an endogenous metalloproteinase inhibitor, i.e. tissue inhibitors of metalloproteinases (TIMPs) [13]. The complex regulation of MMP production by chrondrocytes is affected by several factors, one of which is their response to proinflammatory cytokines, including IL-1β or TNF-α [14-18]. This study was aimed at observing the levels of MMP-1 and MMP-3 and their inhibitors, TIMP-1 and TIMP-2, in CHON-002 induced by IL-1β.
2 Materials and Methods
2.1 Cell Culture
Human long bone cartilage (CHON-002 ATCC ® CRL-2847™) was obtained from Aretha Medika Utama, Biomolecular and Biomedical Research Center, Bandung, Indonesia. The cells were cultured and maintained in Dulbecco's Modified Eagle Media (DMEM, Gibco 11995065) supplemented with 10% (v/v) fetal bovine serum (FBS, Gibco 10270106), 1% anti-biotic and antimycotic (Gibco, 152400620) and 1% geneticin (Gibco, 10151027), and incubated in a humidified atmosphere at a temperature of 37 ºC in 5% CO2 [18].
2.2 Induction with IL-1β
CHON-002 cells (106 cells) were seeded into T-25 flasks and incubated for 2-3 days. Based on a cytotoxicity assay, the viability results for various concentrations showed that the IL-1β was not toxic. Thus, a concentration between 5 and 10 ng/mL is safe and was used for the treatment in this study. The medium was replaced with media containing 5 or 10 ng/mL recombinant IL-1β (Biolegend, 579404). The cells were induced for 5 days [18].
2.3 Cytotoxicity Testing for Viability of CHON-002 Cells after IL-1β Induction
CHON-002 cells at 80% confluency were washed with 1 mL of 1x phosphate buffered saline (PBS, Gibco 14200075) twice. Then, the cells were added with 1 mL of Trypsin-EDTA 0.25% (Gibco, 25200072) and incubated at 37 °C in 5% CO2 for 10 min to remove attached cells. One mililiter of complete medium was then added to stop trypsin digestion. The suspension was centrifuged for 5 min at 1500 rpm. The supernatant was removed and the pellet was homogenized with 1 mL of complete medium. The cells were counted with a haemacytometer after 10x dilution. The cells were plated at 5000 cells/well for 100 µL medium. The cells were added into a 96-well plate and then incubated in 5% CO2 at 37 °C for 24 h. Medium was removed and added with 90% fresh complete medium and 10% of IL-1β. The cells were incubated at 37 °C in 5% CO2 for 24 hours, each well was added with 20 µL of 3-(4,5-dimethylthiazol-2-yl)-5-(3 carboxyme-thoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) (Abcam, ab197010), and the plate was incubated in 5% CO2 for 3 h at 37 °C. Absorbance was measured at 490 nm wavelength using a microplate reader (Thermo Scientific Multiskan Go) [19].
2.4 RNA Extraction and cDNA Synthesis
RNA extraction was done using an Aurum Total RNA Kit (Bio-Rad, 7326820) based on the manufacturer's instructions. Afterwards, the purity of the RNA was measured by spectrophotometry at 260/280 nm with a Thermo Scientific Multiskan Go. Then, the obtained RNA was used for cDNA synthesis using an iScript cDNA synthesis kit (Bio-Rad, 1708890) for 5 min at 25 °C, 30 min at 42 °C, and in the last step for 5 min at a temperature of 85 °C. The product obtained was stored at -20 °C [20].
2.5 mRNA Level Quantification of TIMP-1, TIMP-2, MMP-1, and MMP-3 Genes
Table 1 shows the primer used in this study. Real-time qPCR was done using a PikoReal Real-time PCR System (Thermo Scientific Inc.) with pre-incubation for 5 min at a temperature of 95 °C, continued with 40 cycles of denaturation for 1 min at 95 °C, with the following annealing temperatures: TIMP-1 and TIMP-2 at 50 °C, MMP-1 at 54 °C and MMP-3 at 52 °C for 40 sec, and elongation for 1 min at 72 °C. The reaction mix that was used to perform qPCR was Evagreen Master Mix (Bio-Rad, 1725200) [20].
Table 1 Primer sequences.
| Primer | Forward | Reverse |
|---|---|---|
| β actin | 5'-TCTGGCACCACACCTTCTACAATG-3' | 5'-AGCACAGCCTGGATAGCAACG-3' |
| MMP-1 | 5'-CTGAAGGTGATGAAGCAGCC-3' | 5'-AGTCCAAGAGAATGGCCGAG-3' |
| MMP-3 | 5'-CTCCACGAGCTTGTAGGAAAG-3' | 5'-CACGCCTGAAGGAAGAGATG-3' |
| TIMP-1 | 5'-AGTCAAGACCACCTTATACCA-'3 | 5'-AGTCAACCAGACCACCTTATACCA-3' |
| TIMP-2 | 5'-GGAAGTGGACTCTGGAAACGACATT-3' | 5'-CTCGATGTCGAGAAACTCCTGCTTG-3' |
2.6 Statistical Analysis
The statistical analysis in this study was conducted using SPSS ver. 16.0 with one-way analysis of variance (ANOVA). The analysis was continued with Duncan tests with significancy p < 0.05. The data in this study are presented as mean ± SD.
3 Results
3.1 Viability of CHON-002-IL-1β
The viability of IL-1β-induced CHON-002 cells (CHON-002-IL-1β) was measured using an MTS assay. The assay was based on the formation of purple formazan product from yellow tetrazolium [19]. The cell viability values of the treated cells were compared to control. As can be seen in Table 2, CHON-002- IL-1β on days 1 and 5 showed that the various concentrations of IL-1β (0.1, 1 and 10 ng/mL) and the period of incubation (1 and 5 days) were not toxic. The percentage of viable cells that were given IL-1\(\beta\) showed over 100% live cells and higher IL-1\(\beta\) concentrations showed increased cell viability.
Table 2 Viability of CHON-002 cells treated with different concentrations of IL-1β and periods of incubation.
| Concentration of | Cell viability (%) | |
|---|---|---|
| IL-1β | Day 1 incubation | Day 5 incubation |
| Control | \(100 \pm 13\) | \(100 \pm 25^{a}\) |
| \(0.1 \text{ ng/mL IL-}1\beta\) | \(101 \pm 14\) | \(98 \pm 8^{\mathrm{a}}\) |
| 1 ng/mL IL-1β | \(105 \pm 14\) | \[107 \pm 4^a\] |
| 10 ng/mL IL-1β | \(112 \pm 15\) | \(125 \pm 12^{b}\) |
Note: Data are presented as mean \(\pm\) STD from 3 replications. Superscript letters (a & b) show significant differences between treatment (concentration level of IL-1\(\beta\)) on day 5 of incubation based on Duncan's post hoc test with significancy p < 0.05.
3.2 Characterization of CHON-002-IL-1β
Degenerative changes in OA cartilage are affected by chondrolytic processes that involve MMPs. In this study, the levels of MMP-1 and MMP-3 and its inhibitors, TIMP-1 and TIMP-2, were measured in osteoarthritis-like characteristics of IL-1\(\beta\)-induced CHON-002 cells. The levels of TIMP-1 and TIMP-2 in the IL-1\(\beta\)-induced cells were lower than in control. Meanwhile, MMP-1 and MMP-3 were higher in the IL-1\(\beta\)-induced cells compared to the control cells, which indicates chondrocyte damage (Figure. 1).
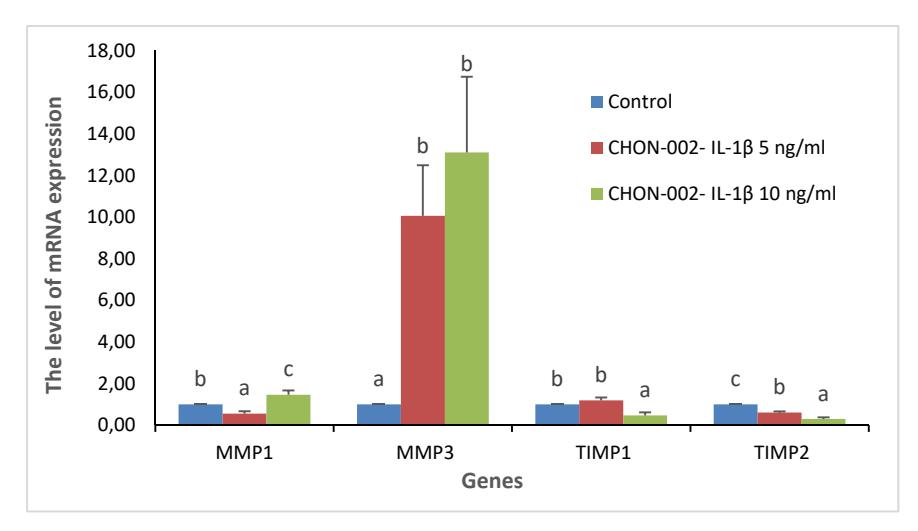
Figure 1 Levels of mRNA expression of MMP-1, MMP-3, TIMP-1 and TIMP-2 genes in IL-1β-induced CHON-002. The data were analyzed with ANOVA and Duncan's post hoc test. The letters (a,b,c) indicate significant differences between treatments (p < 0.05) for each parameter (MMPs and TIMPs).
4 Discussion
In this study, IL-1β was not toxic to the cell line, as indicated by cell viability above 100% after treatment. This result is not consistent with previous research, where IL-1β caused cell death by apoptosis and necrosis. Cell death caused by apoptosis was observed in human chondrocytes in normal OA cartilage after incubation with human IL-1β and after ROS-mediated necrosis was suppressed [21].
In the current study, the levels of MMP-1 and MMP-3 were increased in CHON-002-cells treated with 10 ng/mL IL-1β, while the levels of its inhibitors, TIMP-1 and TIMP-3, were decreased. Several previous studies, both in vivo and in vitro, have shown the role of MMPs in cartilage damage [22-24]. Elevated levels of MMP-1 (collagenase-1) and MMP-3 (stromelysin-1) were documented in osteoarthritic cartilage [25,26] and also found in the synovial fluid of osteoarthristic joints [26,27]. TIMPs are known to block collagenolysis effectively, which indicates a role in the inhibition of MMPs in this process, while aggrecanolysis is blocked by TIMP-3 [28], probably by inhibition of ADAMTS-4 and -5 [29].
Cartilage matrix degradation is considered a result of increased activity of chondrocyte-produced enzymes. The cartilage breakdown involves various kinds of cytokines, while OA chondrocytes are able to synthesize IL-6, IL-1β, TNF-α, and many other enzymes [9,15]. MMP synthesis has been up-regulated by the proinflamatory cytokines TNF-α and IL-1 β by chondrocytes under normal and pathologic conditions [9,18]. Several studies have reported measurable levels of cytokines, such as IL-1β and TNF-α, in synovial fluids found in normal and diseased joints, contributing to expression of MMP by chondrocytes [30]. Referring to previous studies, IL-1β and TNF-α can be produced by human chondrocytes in situ and synthesis of these cytokines by local chondrocytes can be an alternative source of MMP induction in OA cartilage [31].
5 Conclusion
Although apoptotic and necrotic mechanisms can cause cell death, up to 10 ng/mL IL-1β is not toxic to CHON-002 cells. Since this level of IL-1β increased MMP-1 and MMP-3 expression and decreased TIMP-1 and TIMP-2 expression, CHON-002-IL-1β may be a useful model for this aspect of OA.
Acknowledgements
We gratefully acknowledge the financial support of Program Insinas Individu Pratama from the Directorate of Research Empowerment and Development, Ministry of Research, Technology and Higher Education of the Republic of Indonesia (25/INS/PPK/E/E4/2017). We are thankful to Yukko Arinta, Hanna Sari W. Kusuma from Aretha Medika Utama, Biomolecular and Biomedical Research Center, Bandung for their valuable assistance. This study was supported by Aretha Medika Utama, Biomolecular and Biomedical Research Center, Bandung, Indonesia by providing the research laboratory, facilities, and methods.