
1 Introduction
Breast cancer (BC) is one of the most regularly recognized cancer types and is the first cause of female cancer mortality [1]. Metastatis to organs is the main cause of patient mortality [1]. An ideal therapeutic strategy is therefore needed to target metastatic and non-metastatic tumor cells directly, which has to show local activity over the time of treatment [2]. Metastatic tumor cells require invasive and escaping ability in response to immune cells to be able to spread to other organs [3-5]. BC occurrence and its metastatis may also be supported by antitumor immunity [6], immunosurveillance deficiency, natural killer (NK) cell maturation damage, a low number of NK cells and decreased cytotoxic activity in peripheral blood mononuclear cells (PBMCs) [7-10], NK abnormalities [11], low tumor infiltration [12,13], abnormal expression of activating receptors, and overexpression of inhibitory receptors CD158a, CD158b, and NKG2A [10]. Stimulating NK cell functions in combination with other anticancer agents to improve their anticancer activity is a promising cancer therapy [14]. Interleukin-2 (IL-2) can be used to trigger NK cells [15]. Cytoplasmic granules, perforin (Prf1) and granzymes (Gzm) are released by NK cells to suppress cancer, leading to apoptosis [16]. Secretomes such as TNF-α and several effector molecules such as IFN-γ can lead to apoptosis of cancer cells [17]. Another promising anticancer agent is the secretome of mesenchymal stem cells (MSCs) found in conditioned medium (CM), which contains microparticles that mediate therapeutic effects against cancer [18]. MSCs release various factors that can inhibit proliferation of glioma, melanoma, lung cancer, hepatoma, and breast cancer cells [19-22]. Furthermore, MSCs-CM inhibits mammary carcinoma, osteosarcoma, pancreatic, and lung tumor growth [23].
To date, hWJMSC-CM is known to exhibit anticancer activities, yet further development to enhance its potential is still required. An approach suggesting the increase of cytokines such as IL-2 as anticancer treatment may directly induce NK cells to target cancer cells and hWJMSC-CM indirectly. In this research, we aimed to elucidate the properties of IL-2 hWJMSC-CM against BC and NK cell viability, the effect of IL-2 NK cells on BC cell inhibition, NK cell proliferation, and the levels of TNF-α, IFN-γ, perforin, and granzyme, and cytotoxic activity of IL-2 NK against BC cells.
2 Materials and Methods
2.1 Interleukin 2-induced Human Wharton's Jelly Mesenchymal Stem Cells (IL-2 hWJMSCs)
Human Wharton's jelly mesenchymal stem cells (hWJMSCs) at passage 4 (P4) were obtained from our previous study [23]. Surface marker expression and differentiation ability into three different lineages was performed in order to characterize the hWJMSCs [23,24]. The informed consent from all donors and protocol approval of the Institutional Ethics Committee at the Stem Cell and Cancer Institute, Jakarta, Indonesia and the Institutional Ethics Committee collaboration between Maranatha Christian University, Bandung, Indonesia and Immanuel Hospital Bandung, Bandung, Indonesia were obtained to conduct this research. 5 × 105 hWJMSCs were maintained in minimum essential medium-α (MEM-α) (Gibco, 12561056), supplemented with 20% fetal bovine serum (FBS, Gibco 10270106) and 1% of Antibiotic-Antimycotic (Gibco, 1772653) and incubated in 5% CO2 at 37 °C for 24 h. Then, the medium was replaced and the cells were washed with 1 x PBS sterile and supplemented with new growth medium. Next, the cells were treated with 125 and 250 ng/ml of IL-2 (Biolegend, 589106) and incubated in 5% CO2, at 37 °C for 24 h. The IL-2 hWJMSCs were harvested and the medium supernatant was collected, centrifuged at 500 g for four mins at room temperature and filtered through a 0.22 μm Millexe-GV Filter Unit with Durapore (Millipore, SLGV 033 RS). The filtered medium, i.e. conditioned medium of IL-2 hWJMSCs, was stored at -80 °C [23-25].
3 Cytotoxic Activity of IL-2 hWJMSC-CM toward Breast Cancer Cells
The breast cancer cell lines, i.e. MCF-7 (ATCC® HTB22™), were obtained from the Aretha Medika Utama, Biomolecular and Biomedical Research Center, Bandung, Indonesia. Around 5 × 103 cells/well of MCF-7 were maintained in Dulbecco's Modified Eagle's Medium (DMEM; Gibco, 11995065) supplemented with 10% FBS and 1% Antibiotic-Antimycotic. Then, the cells were incubated at 5% CO2, 37 °C for 24 h [23,26,27]. Moreover, various concentrations of IL-2 hWJMSC-CM (30% and 60%) were used to treat the cultured MCF-7, which was incubated for around 24 h. The viability of the cells was counted based on an optimized reagent containing resazurin, which was converted to fluorescent resorufin by viable cells absorbing light at 490 nm in a 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)- 2H-tetra-zolium (MTS) assay (Promega, Madison, WI, USA). The absorbance of the cells was measured at 490 nm using a spectrophotometer (Multiskan Go, Thermo Scientific) [23-26].
3.1 Cytotoxic Activity of IL-2 hWJMSC-CM toward NK Cells
NK92MI cell line (ATCC® CRL2408™) was obtained from Aretha Medika Utama, Biomolecular and Biomedical Research Center, Bandung, Indonesia. 1 × 104 cells/well of NK92MI were plated in a 96-well plate and maintained in RPMI 1640 medium (Gibco, 22400089) supplemented with 10% FBS, 1%
Antibiotic-Antimycotic. Next, the cells were treated with various concentrations of IL-2 hWJMSC-CM (30% and 60%) and then incubated for 24 h. Cell viability in each well was measured by MTS assay, as described in Section 2.2. [23,26].
3.2 Cytoplasmic Granule and Effector Molecule Measurement of hWJMSCs and IL-2 NK Cells in Co-Culture Conditon
NK92MI cells were grown at density \(2 \times 10^6\) cells/well in NK medium containing RPMI 1640, 20% FBS, 1% Antibiotic-Antimycotic. Then, the cells were treated with 10 ng/ml of human recombinant IL-2 and incubated at 5% CO<sub>2</sub>, 37 °C for 96 h, which resulted in IL-2 induced NK (IL-2 NK) cells. The hWJMSCs were cultured at a density of \(1 \times 10^6\) cells/well in MEM-α, supplemented with 10% FBS, 1% Antibiotic-Antimycotic and incubated in 5% CO<sub>2</sub>, 37 °C for 24 h. Furthermore, NK cells at density \(5 \times 10^6\) cells/well were plated in a 24-well plate to achieve a ratio of 1:5 (hWJMSCs: NK = 1:5; hWJMSCs: IL-2 NK = 1:5), containing hWJMSCs in NK medium (RPMI 1640) and MEM-α medium at a ratio of 1:1 (50%:50%) corresponding to optimized conditions (data not shown). The IL-2 NK cells and hWJMSCs coculture were then placed in 5% CO<sub>2</sub> at 37 °C, for 24 h. The resulted medium was preserved at -80 °C for the next assays to measure IFN- \(\gamma\), TNF- \(\alpha\), perforin (Prf1), and granzyme B (GzmB) [25,28,29].
3.3 Cytotoxic activity of CM from Co-culture of hWJMSCs and IL-2 NK toward MCF-7
MCF-7 cells were cultured at a density of \(5 \times 10^3\) cells/well in DMEM medium added with 10% FBS and 1% Antibiotic-Antimycotic and then placed in 5% CO<sub>2</sub> at 37 °C, for 24 h [23,26,27]. Moreover, the cells were treated with (hWJMSCs+NK)-CM, (hWJMSCs+IL-2 NK)-CM at concentrations of 30% or 60% and incubated for 24 h. The viability of the cells was calculated using an MTS assay according to the manufacturer's procedure (Section 2.2) [23,26].
3.4 Apoptosis Assay of CM from Co-culture of hWJMSCs and IL-2 NK toward MCF-7 using Flowcytometer
MCF-7 at density \(1 \times 10^6\) cells/disk were maintained in DMEM medium added with 10% FBS and 1% Antibiotic-Antimycotic and incubated at 37 °C, 5% CO<sub>2</sub> for 24 h. Moreover, the cells were treated using 50% CM of co-culture hWJMSCs and IL-2 induced NK (hWJMSC-CM, (hWJMSCs+NK)-CM, (hWJMSCs+IL12-NK)-CM)) and incubated for 24 h. Next, the cells were washed using phosphate buffered saline (PBS), harvested using 0.25% trypsin EDTA (Gibco, 25200072) and counted using a hemocytometer. Around \(5 \times 10^5\) to \(1 \times 10^6\) counted cells were added with 500 \(\mu\)l of Annexin V binding buffer
(Miltenyi Biotec, 130-092-820), 5 μl of Anti-FITC (Miltenyi Biotec, 130-048- 701) and 5 μl of propidium iodide (Miltenyi Biotec, 130-093-233), and the cells were incubated in darkness at 4 °C. A MACSquant Analyzer 10 (Miltenyi Biotec) was used to analyze the apoptotic percentage of MCF-7 [26].
3.5 Real-time PCR Assay for the Apoptotic Induction of MCF-7 Cell Lines by TNF-α, IFN-γ
Recombinant human TNF-α (Biolegend 570106) and recombinant IFN-γ (Biolegend 570206) were used to determine the apoptosis induction ability toward MCF-7. MCF-7 cells at density 2 × 106 /well were maintained in 500 μl of DMEM supplemented with 10% FBS, 1% Antibiotic-Antimycotic, TNF-α (175, 350 ng/ml), IFN-γ (175, 350 ng/ml) and incubated at 37 °C, 5% CO2 for 24 h [30].
Total RNA from MCF-7 cells was isolated (AurumTM Total RNA Mini Kit, Bio-Rad 732-6820) based on the manufacturer's instructions. The total RNA yield was estimated spectrophotometrically at 260/280 nm (Table 1). The RNA quality was confirmed via electrophoresis, the RNA purity was verified and then the RNA was reverse-transcribed into cDNA (iScript cDNA Synthesis Kit, 170- 8841; Bio-Rad). PCR amplification was carried out using a PikoRealTM Real-Time PCR System (Thermo Scientific Inc.). The qPCR conditions were pre-denaturation at 95 °C for 30 seconds, then 40 cycles of qPCR with denaturation for 5 seconds at 95 °C, annealing for 20 seconds at 58 °C, then elongation for 30 seconds at 72 °C using Sso7d-fusion Polymerase and EvaGreen Dye as detection system (SsoFastTM EvaGreen®Supermix, 172- 5201AP). The data analysis was calculated using Livak formula in Eq. (1):
\[\begin{split} \Delta \text{CT}_1 &\rightarrow \text{target gene expression} \\ &= \text{CT}_{targete gene} - \text{CT}_{house-keeping gene} \\ \Delta \text{CT}_2 &\rightarrow \Delta \text{CT}_{control} \\ &= \text{CT}_{control} - \text{CT}_{house-keeping gene} \\ \Delta \Delta \text{CT} &\rightarrow \Delta \text{CT}_1 - \Delta \text{CT}_2 \\ &= 2 - \Delta \Delta \text{CT} \rightarrow \text{comparison of expression level} \end{split}\]
As internal control, β-actin was included as the house-keeping gene. The primers used for RT-PCR are summarized as: β-actin (forward: 5'-TCT GGC ACC ACA CCT TCT ACA ATG-3', reverse: 5'-AGC ACA GCC TGG ATA GCA ACG-3'), p53 (forward: 5' AGA GTC TAT AGG CCC ACC CC 3', reverse: 5'-GCT CGA CGC TAG GAT CTG AC 3'), Bax (forward: 5'-TGC TTC AGG GTT TCA TCC AG 3', reverse: 5'-GGC GGC AAT CAT CCT CTG
3'), Bcl-2 (forward: 5'-GGT CAT GTG TGT GGA GAG CG -3', reverse: 5'- GGT GCC GGT TCA GGT ACT CA-3') [31].
4 Results
4.1 MSCs Markers and Differentiation
Passage 4 (P4) hWJMSCs strongly expressed CD105, CD73 and CD90 (> 95%) and showed poor expression of CD34, CD45, CD14, CD19 and HLA-II (< 2%) [23]. The cells showed multipotent differentiation into adipocytes, chondrocytes, and osteocytes (data not shown). Therefore, hWJMSCs were confirmed based on the surface marker and differentiation capability [23,24,30].
4.2 Cytotoxic Activity of IL-2 hWJMSC-CMs toward BC Cells
To determine the effect of CM from hWJMSCs treated with IL-2 at concentrations of 125 ng/ml and 250 ng/ml IL-2 hWJMSC-CM against BC cell proliferation, we evaluated the inhibition of IL-2 hWJMSC-CM on MCF-7 at two different concentrations of CM (30% and 60%) (Figure 1).
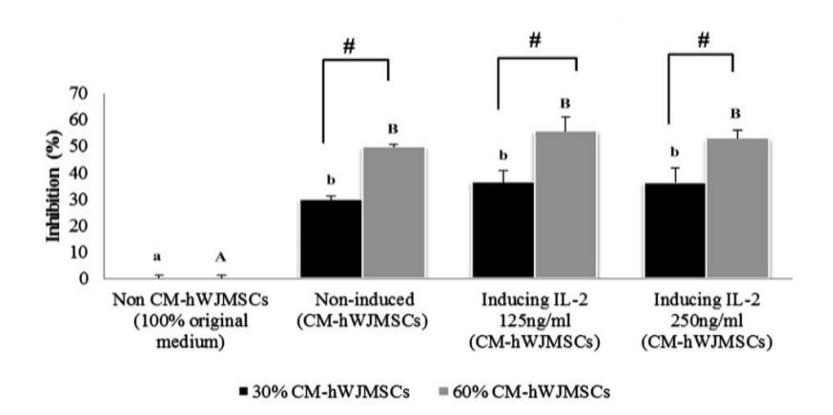
Figure 1 The effect of IL-2 hWJMSC-CM on BC cell growth inhibition.
The data are presented as a histogram of the treatments; the experiment was conducted in triplicate for each treatment. Significant differences between hWJMSC-CM (non-induced, induced by IL-2 125 ng/ml, induced by IL-2 250 ng/ml) and non-hWJMSC-CM (100% MCF-7 medium), based on the Tukey HSD post hoc test (p < 0.05), in 30% and 60% of hWJMSC-CM are indicated by small letters (a,b) and capital letters (A,B), respectively. The # symbol indicates significant differences between the concentrations of 30% and 60% of hWJMSC-CM based on a Student's t-test (p < 0.05). Figure 1 shows that hWJMSC-CM and IL-2 hWJMSC-CM inhibited cell proliferation and had anticancer activities against MCF-7 cells. The higher concentration (60%) of hWJMSC-CM and IL-2 hWJMSC-CM increased the inhibition of BC cells (MCF-7) more compared to the lower concentration (30%) based on a Student's t-test (p < 0.05).
4.3 Effect of IL-2 hWJMSC-CM on NK cell inhibition
IL-2 can activate and improve NK cells. Therefore we induced the hWJMSC-CMs with IL-2 at different concentrations (125 and 250 ng/ml) to see the effect on inhibition of NK cell growth. This experiment was conducted in triplicate for each treatment. The results are shown in Figure 2.
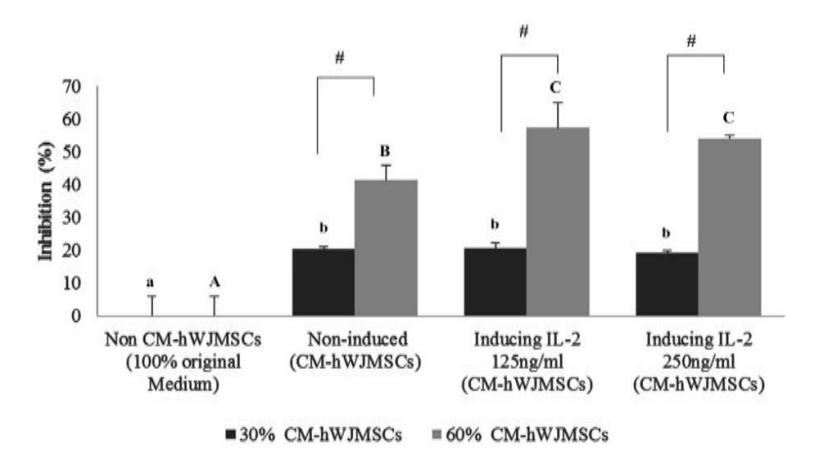
Figure 2 The effect of (IL-2 hWJMSCs)-CM on NK cell growth inhibition.
The data are presented as histograms comparing the treatments. Significant differences between hWJMSC-CM (non-induced, induced by IL-2 125 ng/ml and by IL-2 250 ng/ml) and non-hWJMSC-CM (100% medium of NK cells), based on Tukey's Honest Significant Difference test (p < 0.05), in 30% and 60% of hWJMSC-CM are indicated by small letters (a,b) and capital letters (A,B,C), respectively. The # symbol represents significant differences between concentrations of 30% and 60% of CM-hWJMSCs based on a t-test (p < 0.05). Figure 2 shows that both hWJMSC-CM and (IL-2 hWJMSCs)-CM were toxic against NK cells. The (IL-2 hWJMSCs)-CM was significantly more toxic at higher concentration (60% CM) than at lower concentration (30% CM) of CM, with no significant differences between the different concentrations of IL-2.
4.4 Effect of IL-2 NK cells on the Level of TNF-α, IFN-γ, Perforin, and Granzyme in Co-cultured hWJMSCs and NK Cells
Repairing and degrading damaged tissue is the function of NK cells, in addition to preventing pathogens [32]. A previous study suggested that MSCs have the ability to inhibit the proliferation and improve the functioning of immune cells, showing that MSCs influence the proliferation and cytolytic activities of NK cells [33-35] and the IFN-γ production [36]. NK cells were activated using IL-2 (10 ng/ml) and non-induced-NK cells, which were co-cultured with hWJMSCs. The cells (hWJMSCs:NK cell ratio = 1:5) were assessed for the ability to secrete TNF-α, IFN-γ, perforin, and granzyme B. The effect of IL-2 inducer and NK cell number can be seen in Table 1.
Table 1 Effect of IL-2 inducer and NK cell number on IFN-\(\gamma\), TNF-\(\alpha\), perforin, and granzyme levels in co-cultured hWJMSCs and NK Cells.
| Co-culture hWJMSCs | Concentrations of Effector Molecules Cytoplasmic Granules | |||
|---|---|---|---|---|
| and (IL-2 NK) | IFN-γ (pg/ml) | TNF-α (pg/ml) | Perforin (pg/ml) | Granzyme B (pg/ml) |
| hWJMSC-CM | \(0.94\pm0.19^{a}\) | \(2.64\pm0.47^{a}\) | \(0.00\pm0.00^{a}\) | \(0.00\pm0.00^{a}\) |
| hWJMSC-CM+NK (1:5) | \(22.6\pm1.9^{b}\) | \(8.58\pm0.07^{b}\) | \(90\pm30^{\rm b}\) | \(16.3\pm3.2^{b}\) |
| hWJMSC-CM+IL-2 NK (1:5) | \(23.9 \pm 1.0^{b}\) | \(28.1 \pm 3.0^{\circ}\) | \(180 \pm 10^{c}\) | \(57.33\pm0.58^{c}\) |
The data experiment was performed with 3 replications. The data of concentartions of IFN-\(\gamma\), TNF-\(\alpha\), perforin, and granzyme (pg/ml) are presented as mean \(\pm\) standard deviation. Different small letters in the same column (a,b for IFN-\(\gamma\); a,b,c for TNF-\(\alpha\); a,b,c for perforin; a,b,c for granzyme) indicate significant differences between the treatments based on the Tukey HSD post-hoc test (p < 0.05).
4.5 Effect CM from Co-Culture hWJMSCs and NK Cells towards Cytotoxic Effect of MCF-7
Complex factors that can reduce cancer cell proliferation are secreted by MSCs [21-23]. The hWJMSC-CM can suppress the immunomodulatory system and NK cells and their cytotoxic effect. To increase the cytotoxic effect of hWJMSC and NK cells toward cancer cells, their activation is required. Cytokines such as interleukins can probably be used to directly boost the anticancer property of NK cells and indirectly that of hWJMSC-CM. hWJMSC-CM is known to exhibit anticancer activities because it contains microparticles that mediate therapeutic effects against cancer [18]. The MCF-7 cells were treated with (hWJMSC+NK)-CM, (hWJMSCs+IL-2 NK)-CM at concentrations of 30% or 60% and incubated for 24 h (Table 2).
Based on the cytotoxic data (Table 2), hWJMSC-CM inhibited MCF-7 proliferation and its combination with NK and activated-NK cells increased cytotoxic activity of hWJMSC-CM toward MCF-7.
| CM from a sultaned by IMCC | Proliferation Inhibition toward MCF-7 Cells (%) | ||
|---|---|---|---|
| CM from co-cultured hWJMSCs and IL-2 activated NK Cells | Concentration 30% of CM from co-cultured (hWJMSCs+ IL-2 NK) | Concentration 60% of CM from co-cultured (hWJMSCs+ IL-2 NK) | |
| Non-CM (MCF-7 cells only) | 0.00±6.6 a | 0.00±6.6 a | |
| hWJMSC-CM | 32.66±6.6 b | \(48.0\pm8.7^{\text{ b}}\) | |
| hWJMSC-CM + NK (1:5) | \(42.1\pm3.6^{\text{ bc}}\) | 54.6±8.0 bc | |
| \(hWIMSC_{-}CM + II_{-}2NK(1.5)\) | 61 5+1 3 ° | 68 3+3 8 ° | |
Table 2 Effect of CM from co-cultured hWJMSCs and IL-2 NK cells on MCF-7 proliferation.
The data are presented as mean \(\pm\) standard deviation. The proliferation inhibition was measured in triplicate for each sample. Different superscripts letters in the columns (a,b,bc,c) of 30% and 60% CM concentration of co-culture hWJMSCs and NK cells (number of hWJMSCs: NK = 1:5) indicate significant differences based on Tukey's HSD test (P < 0.05).
4.6 Effect of CM from Co-cultured hWJMSCs and IL-2 NK Cells toward MCF-7 Apoptosis
Secretome components secreted by MSCs, such as chemokines, cytokines, and growth factors, are crucial to maintain the homeostasis and signal transduction with stromal cells (fibroblast, endothel cell, macrophage) [37]. These soluble factors in CM determine the fate of stem cells in in-vitro assays [38]. We measured the apoptosis ability of CM from co-cultured hWJMSC and IL-2 NK cells at a concentration of 50% in normal media towards MCF-7. The (hWJMSCs+IL-2 NK)-CM induced apoptosis of MCF-7 (Table 3, Figure 3).
Table 3 Effect of CM from co-cultured hWJMSCs and IL-2 NK cells toward apoptosis of MCF-7.
| CM-(hWJMSCs + IL-2 NK) | Early Apoptosis (%) | Live (%) | Necrosis (%) | Dead (%) (Late Apoptosis) |
|---|---|---|---|---|
| Non-CM (MCF-7 cells only) | 4.51±0.78 a | 87.1±1.6 ° | 6.74±0.29 b | 1.65±0.78 a |
| hWJMSC-CM | 33.11±0.92 b | 49.3±1.0 b | \(6.70\pm0.28^{\ b}\) | \(10.88\pm0.15^{\ b}\) |
| (hWJMSCs+NK)-CM | 34.7±1.9 b | 53.2±3.8 b | 4.87±0.22 a | 7.3±1.7 b |
| (hWJMSCs+IL-2 NK)-CM | 36.5±2.7 b | 42.47±0.65 a | 5.01±0.87 a | 15.9±1.7 ° |
The data are presented as mean \(\pm\) standard deviation. The number of early apoptosis, live, necrosis, and late apoptosis cells were measured in triplicate for each sample. Different superscript letters in the columns (a,b) of early apoptosis in CM of co-cultured hWJMSCs and IL-2 NK cells and different superscript letters (a,b,c) in the columns of living cells in CM of co-culture hWJMSCs and NK cells, different superscript letters (a,b) in the same column of necrosis in CM of co-culture hWJMSCs and NK cells, different superscript letters (a,b,c) in the same column of late apoptosis in CM of co-culture hWJMSCs and NK cells indicate significant differences based on a Tukey HSD post-hoc comparison (P < 0.05).
Table 3 shows that CM from hWJMSCs, co-cultured hWJMSCs and NK cells, and from co-cultured hWJMSCs and NK cells activated by IL-2 (10 ng/mL) increased the number of early and late apoptotic cells, and reduced the number of live and necrosis MCF-7 cells. The CM from the co-cultured hWJMSCs and IL-2 activated NK cells was more active in inducing early apoptosis and killing
MCF-7 cells (late apoptosis) compared to the CM from co-culture with NK cells without activation, or from hWJMSC-CM alone.
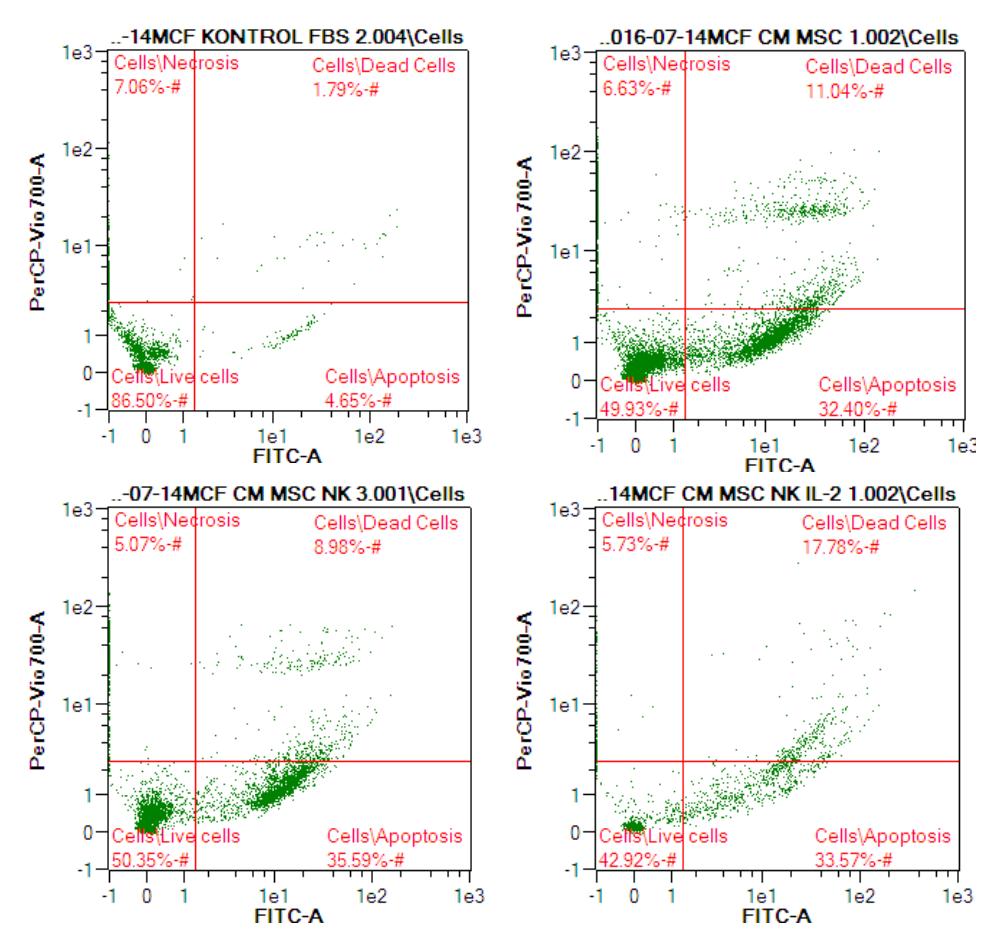
Figure 3 The effect of CM from co-cultured hWJMSCs and IL-2 activated NK cells on apoptosis of MCF-7 cells.
The apoptosis percentage of BC cells treated with CM from co-culture of hWJMSCs and IL-2 activated NK cells was determined by flow cytometry.
- A: Control (MCF-7 cells untreated)
- B: MCF7 treated with hWJMSC-CM
- C: MCF7 treated with (hWJMSCs+NK)-CM,
- D: MCF7 treated with (hWJMSCs+IL12-NK)-CM
Immunotherapy using activated-NK cells has been developed recently [6]. hWJMSC-CM induces apoptosis by upregulating Casp3 and Casp9 as the apoptotic genes and down-regulating the antiapoptotic genes in the human
U251 cell line [39]. The present research was performed to measure proapoptotic and antiapoptotic gene expression of Bax, p53, and Bcl-2 (Table 4).
Table 4 Effect of CM from Co-cultured hWJMSCs and IL-2 NK Cells toward Apoptotic Genes on BC Cells.
| Conditioned Medium (hWJMSCs + IL-2 NK)-CM | Bax | p53 | Bcl-2 |
|---|---|---|---|
| Non-CM (MCF-7 cells only) | 1.00±0.00° | 1.00±0.00° | \(1.00\pm0.00\) |
| hWJMSC-CM | 1.36±0.36 ab | 1.23±0.25 a | \(0.93\pm0.09\) |
| (hWJMSCs+NK)-CM | 1.52±0.17 b | 1.47±0.15 ab | \(0.96\pm0.19\) |
| (hWJMSCs+IL-2 NK)-CM | \(1.46\pm0.16^{b}\) | 1.94±0.44 b | \(0.88\pm0.19\) |
The data are presented as mean \(\pm\) standard deviation. The proapoptotic (Bax, p53) and antiapoptotic genes (Bcl-2) were measured in triplicate for each sample. Different superscript letters (a,ab,) in the same column in 50% CM of co-culture hWJMSCs and IL-2 NK cells indicate significant differences based on a Tukey HSD post-hoc comparison (P < 0.05).
Based on the data in Table 4, NK and activated-NK cells increased apoptotic gene expression (Bax and p53). CM-hWJMSCs and CM from co-culture hWJMSCs and NK cells did not significantly affect antiapoptotic gene expression (Bcl-2).
5 Discussion
CM contains many cytokines, growth factors, putative microvesicles, including (micro) RNA, IL-6, IL-8 [25], tumor growth factor-β1 (TGF-β1), monocyte chemoattractant protein-1 (MCP-1), Regulated upon Activation Normal T Cell Express Sequence (RANTES), collagen type I, fibronectin, secreted protein acidic and rich in cysteine (SPARC), and insulin-like growth factor binding protein-7 (IGFBP-7), which can be used as anticancer agents [36,40].
The ability of IL-2 to improve the anticancer activity of hWJMSC-CM both at CM concentrations of 30% and 60% against MCF-7 was not significantly different (Figure 1). However, the hWJMSC-CM could suppress MCF-7 viability with an inhibition value of around 29-49%. A previous study showed that both CM from normoxic and hypoxic WJMSCs could inhibit cancer cell proliferation, including HeLa, SKOV3, PC3, HSC3, and HepG2 with IC<sub>50</sub> values of 51-74% [24], which is in line with our findings. Another research suggested that a 50% concentration of CM from hWJMSCs repressed breast adenocarcinoma (MDA-MB-231), ovarian carcinoma (TOV-112D), and osteosarcoma (MG-63) cells with 30-60% inhibition [40]. Even though the recombinant IL-2 was unable to improve the anticancer activity of hWJMSC-CM, the cytotoxicity of hWJMSCs against cancer cells can be enhanced using engineering methods such as transfection and transduction. Thus, engineered
hWJMSCs expressing IL-2 can be used as anticancer agents, since hWJMSCs express IL-2 to regulate inflammation and possess anticancer activity via direct killing effects of tumors or positive modulating of the endogenous immune system [41,42].
Furthermore, the hWJMSC-CM and IL-2 hWJMSC-CM inhibited NK cell proliferation (Figure 2). The higher the CM concentration (60% CM), the more toxic the CM was against NK cells. The cytotoxic activity of hWJMSC-CM and (IL-2 hWJMSCs)-CM on NK cells was not significantly different, which indicates that IL-2 induction did not have a major effect on the cytotoxic activity of hWJMSC-CM. A previous study revealed that the hWJMSCs were toxic against NK cells, as MSCs produced PGE2 and TGF-β, which efficiently inhibited activation and suppressed the cytotoxicity effect of downstream resting NK cells toward cancer cells. [43]. Several studies have stated the interaction between NK cells and MSCs, revealing that NK cells killed MSCs and the MSCs strongly inhibited IL-2-induced NK cell proliferation and NK cell cytotoxicity, while CD244 and NKG2D were down-regulated under cell-tocell contact in the interaction between NK and MSCs in the presence of IL-2 or IL-15 or following stimulation by combinations of IL-12/IL-15 and IL-12/1L-18 [33,44-46]. Also, the function of NK cells is impaired by the combination of human adult stem cells (hASCs) and bone marrow mesenchymal stem cells (hBM-MSCs) [47].
There are four mechanisms in activated NK cells to kill targeted cancer cells: releasing cytoplasmic granules (perforin, granzyme), death receptor-induced apoptosis (FasL, TRAIL, TNF-α), effector molecule production (IFN-γ), and antibody-dependent cellular cytotoxicity (ADCC), a result of Fc-γ receptor (FcγR) mediated interaction with NK cells [16,48]. The activation of granzyme and perforin release from NK cells is triggered by binding of FcγR to the Fc domain, causing target tumor cell lysis and Fc-dependent tumor cell phagocytosis [48,49]. NK cells contribute to the tumor death process by monoclonal antibody (mAb) therapy by directing ADCC through Fcg RIIIA (CD16) [50].
The hWJMSC-CM has potential anti-cancer activity toward various cancer cells, such as HeLa, SKOV3, HepG2, PC3, HSC3, A549, HT29, MCF-7 [23, 39]. MSCs inhibit immune system cell proliferation and functions such as NK cell cytotoxicity by suppressing TNF-α and IFN-γ secretion [33-35,51]. However, this research revealed that IL-2 could improve cytotoxicity of NK cells, co-culture IL-2 NK and hWJMSCs by increasing TNF-α, IFN-γ, GzmB and Prf1 secretion (Table 1). ILs (IL-2, IL-12, IL-15, IL-18) and IFNs control NK cell cytotoxic activities [52]. This finding is relevant to another study, which suggested that co-cultured NK cells and human adipocyte stem cells
(hASCs) secrete 40±32.5 pg/ml IFN-γ but co-culture of uninduced NK cells and hASCs does not secrete IFN-γ [47], which is relevant to hWJMSC-CM's cytotoxic effect of killing cancer cells using the apoptosis pathway (Tables 3 & 4).
The production of TNF-α, IFN-γ, Prf1, GzmB increased when the ratio of NK cells was high (Table 1), meaning that the cytotoxic effect of NK cells also increased. This result is relevant to a previous study, where there was interaction between MSCs with both the innate and the adaptive immune system, leading to the activation of some effector functions [53]. The phenotype, cytokine secretion, and proliferation of NK cells were suppressed at lower NK-to-MSCs ratios, which affected the innate immune responses and regulation of adaptive immunity [34]. Moreover, the secretion of IFN-γ was improved when co-cultured hASCs, hBM-MSCs and NK cells were at a ratio of 1:1 for 72 h, as well as co-cultured hASCs and IL-2-expanded NK cells [46,54].
The co-cultured NK cells and AML (K562) cancer cells secreted the highest amount of IFN-γ compared to the co-cultures of NK+hASCs and NK+hBM-MSCs, while the lowest amount of IFN-γ was produced by NK control [46]. The MSCs inhibit the activated NK cells and effector and induction functions such as cytotoxic activity and cytokine production [46]. This is in line with the present research results (Figure 2).
Prf1 mediates cytotoxicity with direct killing by NK cells and indirect killing by secreting death-inducing ligands [55]. Cell growth inhibition and cell death, apoptosis, chromosome condensation and DNA fragmentation, increased Casp-3 activity, and the release of apoptosis-inducing factor (AIF) and cytochrome c from the mitochondria toward cancer cells (HepG2, SK-BR-3, and HeLa) are activated by Prf1 secretion [56].
The surface of the target cells is bounded by Gzm in the concentrationdependent and saturable manners and transported into the cells via endocytosis [57]. The internalization of GzmB into target cells is determined by Prf1 [57]. Apoptosis through the intrinsic mitochondrial pathway is mainly induced by GzmB, either by cleaving Bid or activating Bim, leading to Bak/Bax activation and subsequent signal transduction of Casp-3 activation [58]. GzmB secreted in CM of co-cultured hWJMSCs and NK cells activated apoptosis (Table 1) and also induced the expression of Bax genes (Table 4). Activation of p53 tumor suppressor genes as one of the apoptosis pathways is induced by GzmB to kill cancer cells [59]. Apoptosis in human carcinoma cell lines such as HCT116-3 [60] and MCF-7 [61] is induced using TNF-α, including PKR activation in U937 [62].
IFNγ modulates the pro-apoptotic genes but not pro-necrotic genes in HT-29 as a cell death program through sensitizing the cells [63]. Treatment of IFNγ on a human conjunctival cell line resulted in overexpression of Bax, whereas Bcl-2 and p53 protein levels were not modified. Protecting (Bcl-2, Bcl-Xl) or sensitizing (Bax, bak, bad) causing the cell to undergo apoptosis is controlled by the Bcl-2 protein family [64]. IFNγ-inducible protein 10 (IP10) upregulates p53 and induces Bax expression on HeLa cells [65]. IFN-γ enhances apoptosis by decreasing the mitochodrial membrane potential, upregulating p53 and Bax, down-regulating Bcl-xL, and improving caspase-3 activity [66].
In conclusion, the IL-2 hWJMSC-CM and hWJMSC-CM had anticancer activities against BC cells, where higher CM concentrations increased the inhibition potency. The IL-2 hWJMSC-CM and hWJMSC-CM had toxic activity against NK cells, where higher CM concentrations increased the inhibition potency. A large number of NK and IL-2 inducer cells increased the secretion of TNF-α, IFN-γ, Prf1 and GzmB, which play a role as potent anticancer agents. (hWJMSCs+IL-2 NK)-CM was toxic against BC cells, where a higher concentration of (hWJMSCs+IL-2 NK)-CM increased cytotoxic activities. The (hWJMSCs+IL-2 NK)-CM induced apoptosis in BC cells through increasing apoptotic gene expression (Bax, p53). Further research and preclinical and clinical studies should be pursued prior to therapy application.
6 Acknowledgments
The authors gratefully acknowledge the financial support from the Ministry of Research, Technology and Higher Education of the Republic of Indonesia (Hibah Kompetensi 1598/K4/KM/2017). This research was also supported by the Stem Cell and Cancer Institute, Jakarta, Indonesia and Biomolecular and Biomedical Research Center, Aretha Medika Utama, Bandung, Jawa Barat, Indonesia by providing the laboratory facilities and research methodology. We are thankful to Hanna Sari Widya Kusuma, Annisa Amalia, Fajar Sukma Perdana, Rismawati Laila Qodariah, Wisma Ni Luh Ekayanti from the Biomolecular and Biomedical Research Center, Aretha Medika Utama, Bandung, Jawa Barat, Indonesia, for their valuable assistance.