
1 Introduction
Some bacteria reside underneath plant epidermal cells with no negative implication to the plant's health. These bacteria are called endophytic bacteria. Nearly 300,000 plants are known to host endophytic bacteria, of which the population varies from several to a few hundred strains for each plant species [1]. Endophytic bacteria enter the plant roots through active or passive
Received January 15th, 2019, Revised February 28th , 2021 Accepted for publication March 15th , 2021 Copyright © 2020 Published by ITB Institute for Research and Community Services, ISSN: 2337-5760, DOI: 10.5614/j.math.fund.sci.2021.53.1.2
mechanisms, spread inside the plant and colonize plant organs, such as stem, leaves, flowers, and fruit. They form a complex relationship with their host. Their population densities range from 103-104 CFU per gram of fresh weight [2]. The population of endophytic bacteria in plant organs is influenced by species, age, tillage, and environmental conditions [3,4,5].
Endophytic bacteria have been reported to have a beneficial effect on plant growth and health by producing phytohormones, inducing systemic resistance and fixing free nitrogen in several crops, for example sugarcane [6]. Studies on endophytic bacteria also suggest that these microorganisms indirectly benefit the plant's growth by controlling the growth of pathogenic fungi, for example Fusarium spp. [7,8,9]. The genus Fusarium is considered as one of the main soil-borne pathogens negatively affecting the yield and quality of cultivated plants around the world. Some of their species have a wide range of hosts, such as Fusarium oxysporum. In Indonesia, this pathogen is reported to cause major production losses of vanilla [10], banana [11], and corn [12]. Regarding Fusarium-induced disease in corn, Fusarium infection has been reported to cause a corn production loss of up to 48% [13].
The mechanisms of endophytic bacteria in controlling the growth of fungal pathogens depends on their capabilities. There are several mechanisms of action, such as producing antibiotics, siderophores, and lytic enzymes; competing for substrate and habitat; and activating the plant's systemic resistance [14]. Among bacteria, many members of the genus Bacillus and Pseudomonas have biocontrol ability. They have been reported to suppress fungal pathogen growth via secretion of antibiotics and siderophores, and inducing the plant's systemic resistance [15,16].
Due to the negative effects of the long-term application of chemical fungicides on both agricultural products and the environment, the utilization of endophytic bacteria as potential antifungal-producers emerges as an attractive biotechnological approach. Therefore, in this study we evaluated endophytic bacteria isolated from sweet sorghum (Sorghum bicolor) for their abilities to control a number of well-known pathogenic fungi and identify the mechanisms used to impede fungal growth.
2 Materials and Methods
2.1 Microbial Strains and Media
A total of 78 sweet sorghum endophytes from the stem and root were used in this research. The plants were grown in experimental fields in Cibinong, West Java [17]. Several fungal strains were used for testing, i.e. Sclerotium rolfsii InaCC F5, Fusarium solani InaCC F76, F. oxysporum InaCC F78, and
Colletotrichum gloeosporioides InaCC F264. Trypticase Soy Agar (Difco) medium was used to subculture endophytic bacteria and potato dextrose agar (Difco) medium was used to subculture fungal strains.
2.2 Antagonist Assay of Endophytic Bacteria against Pathogenic Fungi
Antagonist assay was conducted qualitatively by direct confrontation method on malt extract agar (MEA, Oxoid). Each of the fully grown tested fungal strains with a size of 6 mm and in the form of an agar plug was positioned in the middle of an MEA plate. Endophytic bacteria were inoculated near the margins of the agar plate. Four different endophytic bacteria could be tested for their antifungal activities on the same agar plate simultaneously. The bioassay plates were incubated at room temperature for a week. Antagonist assay was performed twice.
2.3 Extraction of Antifungal Compounds
Antifungal-producing endophytic bacteria were inoculated in 100 mL of nutrient broth prepared in a 300-mL Erlenmeyer flask. They were incubated for 5 days in an incubator shaker set to 150 rpm at room temperature. The cultures were then centrifuged for 15 minutes at a speed of 10,000 rpm at 4 °C. The supernatants were extracted using ethyl acetate (1:1). The upper phase in the round bottom flask was evaporated at 40 °C using a rotary evaporator. Sixty µL of the extracts were dropped on a sterilized paper disc and allowed to dry. The paper disc was placed on the agar plates on which fungal strains were growing. They were subjected to incubation for 5 days at room temperature. The antifungal activity was indicated by a clear zone formed around the paper disc. This experiment was further carried out only on the bacteria that previously showed activity against the tested fungi.
2.4 Detection of Non-ribosomal Peptides Synthetases (NRPS) and Polyketides Synthases (PKS) Genes
The primers used to amplify the A domain of NRPS and the KS domain of PKS genes were designed by Amos et al. [18] (Table 1). The polymerase chain reaction mixture to detect the targeted genes contained 12.5 µL of GoTaq Green Master Mix (Promega), 1 µL 0.8 µM of forward primer, 1 µL 0.8 µM of reverse primer, 1.25 µL dimethyl sulfoxide, 8.25 µL of nuclease-free water, and 1 µL template DNA. Amplification of the A domain and the KS domain was performed in an Arktik thermal cycler (Thermo Scientific) using a temperature of 63 °C and 61 °C, respectively, for the annealing process. The amplicons were separated in 2% gel agarose operating at 100 volts. The gel was then stained and examined using a gel documentation system.
Domain Primers Sequences Amplicon (bp) A domain (NRPS) F 5-CGCGCGCATGTACTGGACNGGNGAYYT-3 480 R 5- GGAGTGGCCGCCCARNYBRAARAA-3 KS domain (PKS) F 5-GGCAACGCCTACCACATGCANGGNYT-3 350 R 5-GGTCCGCGGGACGTARTCNARRTC-3
Table 1 Primers used to amplify a domain and KS domain of antifungal producing endophytic bacteria.
2.5 Screening for Cellulase, Chitinase, and Protease Activity
Cellulase assay was carried out on mineral salt medium consisting of 2 g/L KH2PO4, 1.4 g/L (NH4)2SO4, 0.3 g/L MgSO4.5H2O, 0.3 g/L CaCl2, 0.4 g/L yeast extract, 0.005 g/L FeSO4.7H2O, 0.0016 g/L MnSO4, 0.0017 g/L ZnCl2, 0.002 g/L CoCl2, 5 g/L carboxymethyl cellulose-Na, 15 g/L agar in pH 5 [19]. Chitinase assay was carried out on medium consisting of 0.7 g/L K2HPO4, 0.3 g/L KH2PO4, 0.5 g/L MgSO4.5H2O, 0.08 g/L FeSO4.7H2O, 0.001 g/L ZnSO4, 0.001 g/L MnCl2, 20 g/L agar and supplemented with 20 g/L colloidal chitin. Protease activity was evaluated on a basal medium consisting of 1 g/L glucose, 2.5 g/L yeast extract, 20 g/L agar, and supplemented with 10 g/L skim milk casein [20]. Endophytic bacteria were inoculated on each medium and incubated for 2 to 5 days at room temperature. Cellulase test plates were stained with 1 g/L Congo red and further incubated for 15 minutes. Decolorization of Congo red was conducted using 1M NaCl solution. Isolates with a clear zone after incubation were confirmed to produce hydrolytic enzyme. The clear zone index was defined as the ratio between the diameter of the clear zone and the diameter of the isolate.
2.6 Molecular Identification of Endophytic Bacteria
Molecular identification of the collected endophytic bacteria was conducted based on 16S rRNA gene analysis. The polymerase chain reaction mixture contained 12.5 µL of GoTaq Green Master Mix (Promega), 0.5 µL 10 µM of primer 27F (5'-AGAGTTTGATCCTGGCTCAG-3'), 0.5 µL 10 µM of primer 1492R (5'-GGTTACCTTGTTACGACTT-3') [21], 0.5 µL dimethyl sulfoxide, 10 µL of nuclease-free water and 1 µL genomic DNA. Gene amplification was done by pre-denaturation at 94 °C for 90 seconds, denaturation at 94 °C for 30 seconds, renaturation at 50 °C for 30 seconds, elongation at 72 °C for 90 seconds, and final extension at 72 °C for 10 minutes. The process was performed in 35 cycles. The 16S rRNA amplicons were separated on 1% agarose gel and stained using ethidium bromide. The gel was then analyzed under UV light using a gel documentation system.
DNA fragments were sequenced by Macrogen Inc. (South Korea) using 27F and 1492R primers in an ABI 3730xl DNA Analyzer. The sequences were analyzed
using the BioEdit program [22] and then aligned against sequences of type strains in an EzTaxon server [23]. A phylogenetic tree was designed using the MEGA X program [24]. The sequences were aligned using the MUSCLE program [25]. The neighbor-joining method [26] and the Kimura-2 parameter model [27] were chosen to construct the phylogenetic tree. Bootstrap was performed in 1000 replications.
3 Results and Discussions
3.1 Antagonist Assay of Endophytic Bacteria and Their Extracts Against Pathogenic Fungi
Based on antagonist assay using the direct confrontation method, 14 out of the 78 tested isolates showed capability to inhibit fungal growth. The fungal mycelia failed to expand on the agar plates due to bacterial growth, as shown in Fig. 1. Three isolates were active against S. rolfsii and four isolates were active against F. solani with low inhibition activity. Five isolates showed strong inhibition for F. oxysporum mycelia. C. gloeosporioides growth was inhibited by all tested isolates with various levels of inhibition activity. Two isolates, designated as ACNM5 and ACNM6, were able to inhibit all four fungal strains (Table 2).
Figure 1 Antagonist assay of sorghum endophytes: (a) ACNM5 against S. rolfsii InaCC F5, (b) ACNM6 against F. solani InaCC F76, (c) ACNM5 and ACNM6 against F. oxysporum InaCC F78, (d) ATNL2 and ATNM3 against C. gloeosporioides InaCC F264. Mycelium growth was inhibited by bacterial strains.
The four fungal species used in this study are known to be pathogenic for various plants. Although it has been reported that sweet sorghum is mostly damaged by Sphacelotecha sorghi, Sphacelotecha reiliana, and Tolyposporium enherenbergii, other fungi also pose a major obstacle to sorghum production. These pathogenic fungi, including Aspergillus, Fusarium, Rhizoctonia, Penicillium, Sclerotium, and Curvularia, have been identified as seed-borne pathogens for sorghum [28]. The use of C. gloeosporioides, which was commonly related to anthracnose disease on chili in this research, was used to evaluate the efficacy of the collected endophytic bacteria against a wide range of causative agents of plant disease.
Further analysis using bacterial extracts revealed that most of their secondary metabolites were unable to inhibit fungal growth. Only one isolate, designated as ACNM4, retained its activity against fungal strains. The solvent used to extract bioactive compounds may influence the successfulness of the extraction process. Some bioactive compounds may effectively be extracted using a polar solvent and others may not. The concentration of obtained metabolites also plays an important role in their activity to inhibit fungal growth.
| Table 2 Antifungal activity of endophytic bacteria against several fungal | ||||
|---|---|---|---|---|
| strains. |
| Bacterial Cells | Ethyl Acetate Extracts | |||||||
|---|---|---|---|---|---|---|---|---|
| Isolates | InaCC | InaCC | InaCC | InaCC | InaCC | InaCC | InaCC | InaCC |
| F5 | F76 | F78 | F264 | F5 | F76 | F78 | F264 | |
| BTIH1 | - | - | - | + | * | * | * | - |
| BTIL3 | - | - | - | + | * | * | * | - |
| BTIL4 | - | - | - | + | * | * | * | - |
| BTIL5 | - | - | - | + | * | * | * | - |
| BTIL6 | - | - | - | + | * | * | * | - |
| BTIL7 | - | - | - | + | * | * | * | - |
| ACNM4 | + | - | ++ | ++ | + | * | + | + |
| ACNM5 | ++ | + | ++ | ++ | - | - | - | - |
| ACNM6 | ++ | + | ++ | ++ | - | - | - | - |
| ACIL1 | - | - | - | + | * | * | * | - |
| ATNL2 | - | - | - | ++ | * | * | * | - |
| ATNM3 | - | - | ++ | ++ | * | * | - | - |
| ATIM2 | - | + | ++ | ++ | * | - | - | - |
| ATNM4 | - | + | - | ++ | * | - | * | - |
Note: + = tested isolate showing antifungal activity, additional mark indicating stronger activity; - = tested isolate showing no antifungal activity; * = test was not performed as it showed no activity using bacterial cells.
3.2 Detection of NRPS and PKS Genes
The production of secondary metabolites from microorganisms is mainly conducted via non-ribosomal peptides synthetases (NRPS) and polyketides synthases (PKS) pathways. They are multidomain enzymes consisting of peptidepeptidyl carrier protein (PCP), adenylation (A) domain, and condensation (C) domain for NRPS and acyltransferase (AT) domain, acyl carrier protein (ACP) domain, and ketosynthase (KS) domain for PKS [29,30]. The primers employed in this study targeted the A domain of NRPS and the KS domain of PKS, specifically PKS type II. Based on detection of genes involved in bioactive compound production, four isolates appeared to harbor A domain (Fig. 2), i.e. ACIL1, ACNM4, ACNM6, and ATNM4. PCR products were observed in ~480 bp on stained agarose gel. This result suggests that the antifungal compounds were produced via NRPS pathway.
Non-ribosomal peptides with antimicrobial properties are highly detected in bacteria such as Proteobacteria, Actinobacteria, Firmicutes, and Cyanobacteria groups [31]. An eminent example of an antimicrobial compound synthesized via NRPS is penicillin, produced by a group of fungi [32]. Tyrocidine, cephalosporine, daptomycin, vancomycin, quinoxaline, capreomycin, bleomycin, and cyclosporin production is also coded by NRPS genes. These natural products have antibacterial, antifungal, and even anti-tumor capabilities [33].

Figure 2 Gel electrophoresis of A domain amplified by PCR. A domain of isolate ACIL1, ACNM4, ACNM6, and ATNM4 were detected at ~480 bp. Well: (M) Marker DNA ladder 100 bp, (1) BTIL5, (2) BTIL7, (3) BTIL4, (4) BTIL3, (5) BTIL6, (6) ACIL1, (7) ACNM4, (8) ACNM6, (9) ATIM2, (10) BTIHI, (11) ATNM3, (12) ACNM5, (13) ATNM4, (14) ATNL2, and (C) control = nucleasefree water.
The KS domain was not detected in any endophytic bacteria, indicating that the antifungal compounds were not synthesized via PKS pathway. However, it is possible that the primers employed in this study were unable to amplify the existing KS domain of the plant endophytic bacteria. The primers were originally designed to amplify the KS domain of soil bacteria in European, Antarctic, and Cuban soils. Furthermore, antifungal activity of the isolates may have been induced by other mechanisms.
3.3 Screening for Cellulase, Chitinase, and Protease Activity
Fungal cell walls are constituted by chitin, β-glucan, mannan, and protein in most filamentous fungi. However, the composition of those cell wall constituents frequently varies among species of fungi. The mechanical strength of the cell wall influences their shape and integrity. The cell wall performs a wide range of important roles during their interaction with environment [34].
Especially chitin is an important element in the fungal cell wall structure as it gives rigidity to hold out against physical as well as chemical attacks encountered by the fungal cell wall [35]. Therefore, breaking these cell wall components is an essential mechanism in controlling fungal survival.
Hydrolytic enzymes such as chitinase and proteinase produced by bacterial groups have long been reported of being able to cut the glycosidic and peptide bonds of filamentous fungi. Crude chitinase from Serratia marcescens has been reported to cause the lysis of hyphal tips of pathogenic fungi S. rolfsii up to 60% [36]. This biological control activity is also shown by Pseudomonas stutzeri against F. solani, involving bacterial hydrolytic enzyme, chitinase, and laminarinase [37].
In this research, assay on 14 endophytic bacteria with antifungal activity revealed that 10 isolates were able to produce hydrolytic enzymes (Table 3). Only two isolates, designated as BTIL3 and BTIL6, showed cellulase activity on mineral salt medium containing carboxy methyl cellulose. Cellulose is not the main constituent of fungal cell walls. In fact, only a small group of fungi harbor this polysaccharide in their cell wall. These two isolates also produced chitinase with a lytic index of 1.67 and 2.13, respectively. This result suggests that chitin cell wall destruction is not the main mechanism for inhibiting fungal growth. However, some pathogenic fungi, for example, Cladosporium fulvum have evolved chitin-binding effector to defend themselves from hydrolysis and detection by chitinase produced by their plant host [38]. Such a mechanism is a sophisticated response to host defense as well as antagonist microbes.
Table 3 Biocatalyst activity of antifungal-producing endophytic bacteria.
| Isolates Sources | Hydrolytic Enzyme (Index) | Isolates Sources | Hydrolytic Enzyme (Index) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Ce | Ch | Pr | Cl | Ch | Pr | ||||
| BTIHI | Stem | - | - | - | ACNM5 | Root | - | - | 2.92 |
| BTIL3 | Stem | 3.00 | 1.67 | 2.25 | ACNM6 | Root | - | - | 2.60 |
| BTIL4 | Stem | - | - | - | ACIL1 | Root | - | - | - |
| BTIL5 | Stem | - | - | 3.83 | ATNL2 | Root | - | - | 2.11 |
| BTIL6 | Stem | 2.50 | 2.13 | - | ATNM3 | Root | - | - | - |
| BTIL7 | Stem | - | - | 1.40 | ATIM2 | Root | - | - | 3.32 |
| ACNM4 | Root | - | - | 2.42 | ATNM4 | Root | - | - | 3.00 |
Note: Cl = cellulase; Ch = Chitinase; Pr = Protease
Sixty-four percent of tested isolates were protease producers. Their protease indices range from 1.40 to 3.83. The majority of structural proteins found in fungal cell walls are glycosylated and contain mannose, called mannoproteins. Based on the number of isolates capable of producing protease compared to the antagonism assay result, the mechanism employed by endophytic bacteria to inhibit fungal mycelial growth mostly involves their proteolytic activity.
In addition, this result showed that endophytic bacteria isolated from sorghum roots tend to have better antifungal activity. This is probably due to the high competition among microbes that reside around the plant root area in the soil. The soil ecosystem is considered to consist of the greatest number of genetic variations existing on earth. According to estimations, one gram of soil contains as many as \(10^{10}\) viruses, up to \(10^{10}\) bacteria, \(10^6\) fungi, \(10^6\) algae, \(10^5\) protozoa, and \(10^2\) nematodes [39]. This requires a strong technique to defend themselves in order to survive, for example by producing antifungal compounds. Endophytic bacteria producing antifungal compounds can protect their host from pathogenic fungi infection.
3.4 Molecular Identification
The analysis results of the nucleotide sequences of the antifungal-producing endophytic bacteria are shown in Table 4. The isolates showed high similarities with some species in the genera Bacillus, Burkholderia, Staphylococcus and a group of Actinobacteria. The phylogenetic relationships of these antifungal-producing endophytic bacteria are shown in Fig. 3. Endophytic Bacillus and Staphylococcus have also been isolated from sorghum in Uruguay. They showed a plant growth promotion effect on sweet sorghum by producing either protease or exo-cellulase and siderophore regarding their potential as biocontrol agents [40].
| Table 4 Identities of | antifungal-producing | endophytic | bacteria | Based | on | 16S |
|---|---|---|---|---|---|---|
| rRNA gene analysis. |
| Isolates | InaCC Number | Close Relative | Sequence Similarity (%) | Sequence Length (bp) | |
|---|---|---|---|---|---|
| BTIH1 | InaCC B1274 | Paracoccus chinensis | 97.45 | 1298 | |
| BTIL3 | InaCC B1268 | Bacillus cereus | 99.93 | 1376 | |
| BTIL4 | InaCC B1267 | Brachybacterium paraconglomeratum | 99.85 | 1334 | |
| BTIL5 | InaCC B1265 | Janibacter indicus | 99.70 | 1325 | |
| BTIL6 | InaCC B1269 | Bacillus cereus | 99.41 | 1360 | |
| BTIL7 | InaCC B1266 | Microbacterium testaceum | 99.55 | 1334 | |
| ACNM4 | InaCC B1271 | Burkholderia cepacia | 99.41 | 1351 | |
| ACNM5 | InaCC B1276 | Burkholderia cepacia | 99.70 | 1324 | |
| ACNM6 | InaCC B1272 | Burkholderia cepacia | 99.56 | 1367 | |
| ACIL1 | InaCC B1270 | Cupriavidus metallidurans | 98.80 | 1337 | |
| ATNL2 | InaCC B1278 | Bacillus aryabhattai | 99.86 | 1382 | |
| ATNM3 | InaCC B1275 | Staphylococcus lugdunensis | 99.49 | 1361 | |
| ATIM2 | InaCC B1273 | Burkholderia cepacia | 99.92 | 1361 | |
| ATNM4 | InaCC B1277 | Burkholderia cepacia | 99.85 | 1344 | |
Five isolates identified as Burkholderia were capable of inhibiting at least two fungal strains. Their overall antagonist performance was better than that of the other bacteria tested in this study. All of their species were isolated from root
samples. Endohytic Burkholderia can also be found in the stems of sugarcane and maize [41]. They influence plant growth through nitrogen fixation. Three identified isolates belong to the Actinobacteria group, namely Brachybacterium, Janibacter, and Microbacterium. They were isolated from stem samples and like other stem endophytes they were only active against C. gloeosporioides. For decades, Actinobacteria have been known to produce various secondary metabolites with antimicrobial activity, especially Streptomyces. Endophytes have also been reported to have strong activity against pathogenic fungi, C. musae and F. oxysporum [42].
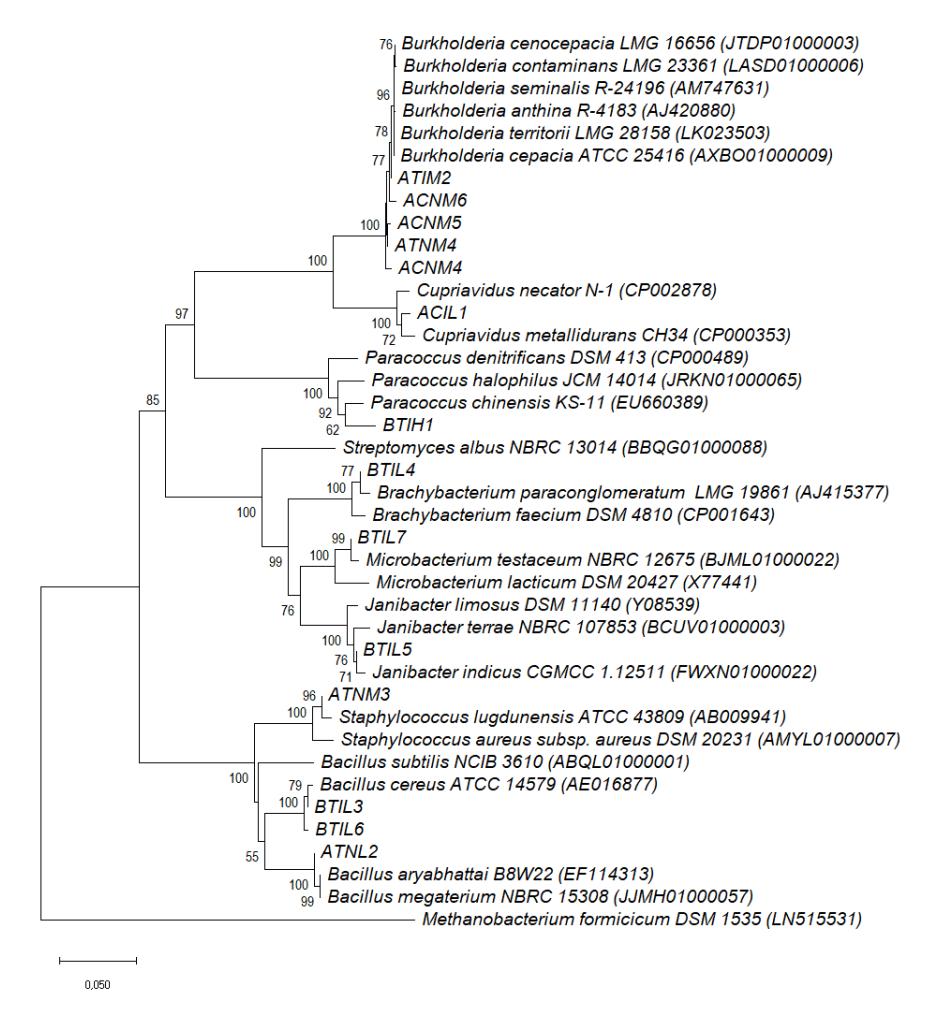
Figure 3 The evolutionary relationship of sweet sorghum endophytes with antifungal capability as constructed on the basis of their 16S rDNA sequences. Bootstrap values are shown at each branch point as percentage. Only values greater than 50% are shown.
Finally, this research demonstrated the important role of endophytic bacteria in the sorghum plant as antifungal producers. Endophytic bacteria can indirectly promote plant growth with their ability to combat plant pathogens. They harbor genes involved in bioactive compound synthesis and are thus capable of producing potential secondary metabolites. The ability to inhibit fungal growth is also supported by their ability to secrete hydrolytic enzymes that play a role in destroying fungal cell walls. Study on their colonization in plants is expected to be done in the near future as a basic step to utilize these endophytic bacteria as biocontrol agents.
4 Conclusion
The isolation methods for stems and roots of sweet sorghum obtained a total of 14 endophytic bacteria showing antifungal activity against Sclerotium rolfsii, Fusarium solani, Fusarium oxysporum, and Colletotrichum gloeosporioides. Two isolates, designated as ACNM5 and ACNM6, are potential wide-range biocontrol agents due to their ability to inhibit all tested fungal strains. Determination of mechanisms of fungal inhibition by detecting specific primers involved in the biosynthesis of natural products, extracting chemical compounds, and screening for lytic enzymes revealed that four isolates, designated as ACIL1, ACNM4, ACNM6, and ATNM4, contained NRPS gene sequences. However, only the chemical compound ACNM4 extracted by ethyl acetate was able to inhibit mycelial growth. Hence, this chemical compound was suggested to be synthesized via NRPS pathway. Based on lytic enzymatic tests, these endophytic bacteria were able to produce cellulose, chitinase, or protease, where the last enzyme was produced by the majority of the tested bacteria. Molecular identification showed that most of these endophytic bacteria with antifungal activity belong to the Burkholderia group.
Acknowledgments
This work was funded by JICA-JST within the framework of the SATREPS Project entitled 'Revegetation of Alang-alang (Imperata cylindrica) Field Combined With Sustainable Production and Utilization of Biomass for Food and Energy Solutions' and the Research Competitive Grant Project of LIPI 2018. The authors would like to thank Rinatu Siswi, Gita Azizah Putri, Dinihari Indah Kusumawati, Mia Kusmiati, Ismu Purnaningsih for their technical assistance, and Muhammad Ilyas for providing the fungal strains employed in this research.