
PENDAHULUAN
Industri tekstil dan produk tekstil (TPT) di Indonesia diyakini mampu memberikan kontribusi signifikan terhadap perekonomian (Dwipayani dan Notodarmodjo, 2013) penerimaan devisa dari kegiatan ekspor. Tercatat sepanjang Januari-Juni 2018 nilai pengapalan produk TPT Indonesia sudah mencapai USD 3,73 miliar, dengan peningkatan 3,86% per tahunnya (Kementerian Perindustrian, 2018). Namun, terlepas dari peranannya sebagai komoditi ekspor yang diandalkan, nyatanya industri tekstil ini menimbulkan masalah yang serius bagi lingkungan terutama masalah yang diakibatkan oleh limbah cair yang dihasilkan (Ayu dan Roosmini, 2014).
Sekitar 2-50% residu dari pewarna sintetik yang digunakan akan tersuspensi pada limbah cair tersebut (Sheth & Dave, 2009). Dalam kasus pewarna reaktif, karena tingkat fiksasi yang rendah terhadap serat, sebanyak 50% dari konsentrasi pewarna awal yang digunakan masih tertinggal pada air pencucian dalam bentuk terhidrolisis dan tidak lagi memiliki afinitas terhadap serat, sehingga tidak dapat digunakan kembali (Watanapokasin dkk., 2008).
Pada jenis pewarna reaktif, azo dan antrakuinon adalah kromofor utama yang sering digunakan (Erkurt dkk., 2009). Saat ini, parameter warna belum dimasukan dalam baku mutu limbah cair industri tekstil. Namun, keberadaan pewarna reaktif di perairan menjadi permasalahan serius pada waste water treatment plan (WWTP) dikarenakan intensitas warna, sifatnya yang bio-rekalsitran serta potensi toksisitasnya terhadap lingkungan (Jadhav dkk., 2010).
Pada studi toksisitas lingkungan menggunakan organisme uji bakteri, ditemukan fakta bahwa pewarna azo Orange 2 (250-500 mg/L) dapat menghambat pertumbuhan P. syringae, E. coli, B. subtilis, M. luteus, dan M. aeruginosa. Bakteri-bakteri tersebut dipilih sebagai organisme yang umum ditemui di lingkungan (Legerská dkk., 2016). Sedangkan, studi toksisitas lain dari pewarna Reactive black 5 yang diujikan terhadap S. meliloti (bakteri
nitrifikasi), menunjukan adanya penurunan viabilitas sel pada konsentrasi pewarna yang semakin tinggi, EC50 yang terukur yaitu pada konsentrasi 164 mg/L (Enayatizamir dkk., 2011).
Penyisihan pewarna dengan metode biologis secara umum banyak dipertimbangkan karena ramah lingkungan dan mengarah pada proses mineralisasi polutan organik secara menyeluruh dengan biaya yang relatif ekonomis. Saat ini, banyak peneliti memanfaatkan kemampuan jamur terutama jamur pelapuk putih, untuk penyisihan berbagai polutan karena sistem ekstraseluler enzimnya yang non-spesifik. Enzim oksidatif ekstraseluler yang di ekskresikan Jamur Pelapuk Putih terdiri dari Mangan Peroksidase (MnP), Lakase, dan Lignin Peroxidase (LiP). Ditinjau dari beberapa hasil penelitian sebelumnya, menyatakan bahwa diantara berbagai jenis enzim, lakase menunjukan potensi yang besar untuk diaplikasikan dalam bidang bioteknologi dan lingkungan (Rivera-Hoyos dkk., 2013; Senthivelan dkk., 2016). Beberapa penelitian telah dilakukan terhadap kelompok jamur yang mampu menyisihkan pewarna antara lain Phanerochaete chrysosporium (Senthilkumar dkk., 2014), Pleurotus ostreatus (Erkurt dkk., 2007), Trametes sp. (Wang dkk., 2018), Marasmiellus palmivorus (Centele dkk., 2017), dan Ganoderma sp. BTA1 (Martina, 2005). Namun, penelitian mengenai kemampuan penyisihan warna oleh jamur pelapuk putih tersebut masih terbatas pada satu jenis pewarna oleh masing-masing isolat jamur.
Penelitian ini bertujuan untuk mengidentifikasi potensi berbagai jenis Jamur Pelapuk Putih untuk menyisihkan pewarna tekstil jenis antrakuinon dan azo, baik yang memiliki ikatan monoazo maupun diazo, dengan menggunakan berbagai isolat jamur pelapuk putih yaitu: Pleurotus ostreatus dan Trametes versicolor sebagai salah satu jenis jamur yang dikenal baik dalam memproduksi enzim lakase (Kocyigit dkk., 2012); Marasmiellus palmivorus sebagai jamur yang banyak tumbuh di indonesia; serta Ganoderma sp., dan Phanerochaete chrysosporium yang telah banyak disebutkan dalam berbagai hasil penelitian (Senthilkumar dkk., 2014; Dias dkk., 2003). Kelima jenis jamur pelapuk putih tersebut kemudian diidentifikasi efisiensi penyisihannya terhadap pewarna Reactive Blue 4 (RB4), Reactive black 5 (RB5), Reactive red 120 (RR120), dan Reactive orange 16 (RO16).
METODOLOGI
Isolat jamur dan Reagen
Isolat fungi yang digunakan adalah Pleurotus ostreatus, Phanerochaete chrysosporium, Marasmiellus palmivorus, dan Ganoderma sp. yang didapatkan dari Laboratorium Mikologi, Pusat Ilmu Hayati, SITH ITB. Sementara Trametes versicolor didapatkan dari Indonesian
Culture Collection, Cibinong – Jawa Barat. Isolat fungi tersebut ditumbuhkan pada medium PDA, kemudian diinkubasi pada suhu ruangan selama 7 hari.
Secara terpisah, media cair Kirk termodifikasi dibuat. Media Kirk dibuat dengan mencampurkan 9 jenis reagen, yaitu glukosa 10 g/L, KH2PO4 1,7 g/L, MgSO4 0,02 g/L, yeast extract 0,3 g/L, CuSO4.7H2O 0,01 g/L, Na2MoO4 0,1 g/L, MnSO4.H2O 0,1 g/L, ZnSO4.7H2O 0,006 g/L, Fe2(SO4)3 0,007 g/L (Hanung dkk., 2013). Seluruh reagen yang digunakan merupakan Analytical grade.
Screening Penyisihan Pewarna pada Medium PDA
Media PDA ditambahkan dengan masing-masing pewarna (RB4, RB5, RR120 dan RO16). Medium tersebut digunakan untuk pengamatan penyisihan warna oleh isolat jamur pelapuk putih yang digunakan. Dengan menggunakan inoculating punch, isolat fungi diinokulasikan dengan diameter 5mm pada medium PDA yang telah diberi warna dengan konsentrasi 150 mg/L. kultur tersebut kemudian di inkubasi selama 24 hari pada suhu ruang (26 0C) untuk diamati zona bening yang terbentuk di sisi bawah petri dish kultur dan diamati setiap 24 jam, serta diekspresikan dalam cm/hari (Rathnan dkk., 2013).
Analisis Kurva Aktivitas enzimatik T. versicolor dan P. ostreatus
Aktivitas enzimatis 2 isolat terbaik yang didapat dari percobaan skala in vitro dilakukan analisis untuk mengetahui enzim yang dominan diproduksi oleh jamur pelapuk putih yang digunakan. Enzim yang dianalisis berupa lakase, Mnp dan Lip selama 12 hari untuk melihat waktu optimal diproduksinya enzim dan enzim yang mendominasi. Erlenmeyer dengan volume 1 liter disiapkan dengan 750 ml medium kirk dan sebanyak 5 mL di inokulasikan isolat jamur yang sudah ditumbuhkan dalam medium PDB selama 7 hari (Centele dkk., 2017). Setiap 24 jam diamati aktivitas enzim (lakase, Mnp dan Lip), kemudian didapatkan 1 isolat dengan aktivitas terbaik, serta 1 enzim yang paling dominan diproduksi jamur dalam medium Kirk (Patrick dkk., 2011; Hafilah, 2017).
Penyisihan pewarna sintetis dalam Submerged fermentation form (SFF)
Isolat terbaik kemudian ditumbuhkan dengan cara yang sama pada percobaan analisis kurva aktivitas enzimatis (Patrick dkk., 2011; Hafilah, 2017). Namun, kultur stok yang digunakan sebesar 5% (v/v) dari volume total dan waktu inkubasinya selama 5 hari. Setiap 24 jam diamati aktivitas enzim, penyisihan warna, Biomassa serta pH. Seluruh perlakuan dilakukan dengan 3 kali pengulangan (triplo). Masing-masing sampel sebelum dan sesudah
diolah, kemudian diukur perubahan panjang gelombang maksimum dengan rentang 200-800 nm untuk mengetahui mekanisme yang dominan yaitu biosorpsi atau biotransformasi (Zheng dkk, 2011; Patrick dkk., 2011; Erkurt dkk., 2007).
Pengukuran Efisensi Penyisihan Warna dan Aktivitas Lakase
Sampel media cair yang dipisahkan dengan miselia melalui proses sentrifugasi 3500 rpm selama 10 menit. Pengamatan penyisihan warna dilakukan dengan menggunakan Spektrofotometer Optizen 2120UV pada panjang gelombang maksimum masing-masing pewarna reaktif (Jayapal dkk., 2018).
Aktivitas lakase diuji dengan ABTS atau 2,2'-azinio-bis-(3-ethylbenzhiazoline-6 sulphonic acid) sebagai substrat uji (Johannes dkk., 2006; Risdianto, 2007; Hanung dkk., 2013; Margot dkk., 2013; Trejo dkk., 2015). Aktivitas Manganese peroksidase (MnP) diuji berdasarkan estimasi penghilangan substrat fenol red (Kuwahara dkk.,1984; Hafilah, 2017). Sedangkan, Aktivitas Lignin Peroksidase (LiP) ditentukan berdasarkan reaksi demetilasi metilen biru (Magalhães dkk.,1996; Hafilah, 2017).
Uji stabilisasi pH dan suhu ekstrak kasar enzim lakase
Pengaruh pH pada aktivitas laccase ditentukan menggunakan buffer asam sitrat-Na2HPO4. ABTS digunakan sebagai substrat (Wang dkk., 2018). Dalam uji stabilitas pH, ekstrak kasar enzim lakase dipreinkubasi pada suhu 4 0C dan selanjutnya, aktivitas diukur menggunakan uji standar (substrat ABTS) dalam asam sitrat-buffer Na2HPO4 (50 mM) pada pH 2,2 hingga 8,0 dengan rentang pH 0,2 (Ling dkk., 2015). Thermo stabilitas diujikan setelah mendapatkan kondisi pH optimum, enzim di pre inkubasi pada suhu 4 0C kemudian, diinkubasi pada berbagai suhu berkisar 20 0C–80 0C dengan rentang 5 0C selama 60 menit dan dilakukan pengujian aktivitas lakase.
Penyisihan warna dengan Ekstrak kasar enzim lakase dan uji toksisitas hasil metabolitnya
Penyisihan warna dengan menggunakan crude enzim lakase dilakukan pada pH dan suhu optimum enzim, sebanyak 1,5 ml larutan stok warna 2000 ppm dicampurkan dengan 18,5 ml ekstrak kasar enzim sehingga konsentrasi warna menjadi 150 ppm. Kemudian sampel diinkubasi selama 60 menit dan diukur persen penyisihan warnanya. Sampel dengan persen penyisihan warna terbaik kemudian dilakukan analisis lanjutan menggunakan instrument FTIR untuk mengetahui biotransformasi gugus fungsi pewarna sebelum dan sesudah diolah (Zheng dkk., 2011).
Koloni Bacillus sp. ditumbuhkan pada medium nutrient broth (NB) selama 24 jam. Toksisitas RB4 dan produk hasil biotransformasinya dianalisis dengan menginokulasikan 1% (v/v) dari kultur yang sudah berumur 24 jam kedalam tabung tes yang mengandung NB dan RB4 sebelum diolah (150 ppm) atau RB4 yang sudah terolah dalam volume total 20 mL. Tabung tes kemudian di inkubasi pada suhu 26 0C dan diagitasi 140 rpm selama 24 jam. Pengencerah bertingkat kemudian disiapkan (aquades steril) dan 1 mL masing-masing percobaan ditambahkan kedalam petri yang sudah diberikan NA untuk selanjutnya diamati jumlah koloni dengan metode Total plate counter (Legerská dkk., 2016; Enayatizamir dkk., 2011).
HASIL DAN PEMBAHASAN
Zona penyisihan warna isolat fungi pada medium PDA yang diberi pewarna reaktif
Pada Gambar 1 Dari keempat warna yang digunakan, hanya isolate P. ostreatus dan T. versicolor yang dapat menyisihkan seluruh warna yaitu RO16, RR120, RB4 dan RB5. Sehingga keduanya sangat potensial untuk menyisihkan pewarna tekstil reaktif dan dapat dilakukan analisis lanjutan untuk mengetahui mekanisme penyisihan yang terjadi.
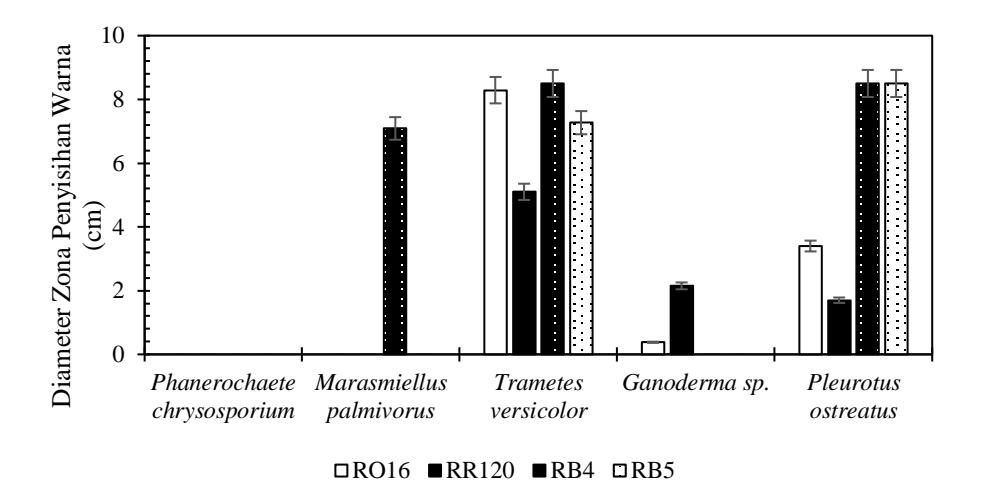
Gambar 1. Grafik Zona penyisihan warna oleh JPP pada SFF
Analisis Kurva Aktivitas enzimatik T. versicolor dan P. ostreatus
Lignin Modified Enzyme (LME) dihasilkan oleh jamur pelapuk putih sebagai metabolit sekunder (Erkurt dkk., 2009), sehingga apabila ingin mendapatkan hasil enzim pendegradasi
warna maka pertumbuhan miselia jamur harus dioptimalkan terlebih dahulu. Setelah kultur stok di inkubasi selama 12 hari, dilakukan pengukuran kadar protein serta aktivitas enzimatis untuk mendapatkan gambaran apakah Isolat yang digunakan mampu menghasilkan enzim yang diharapkan. Medium cair yang digunakan mengandung glukosa sebagai sumber karbon sederhana yang dapat dimanfaatkan oleh jamur secara langsung. Namun, kehadiran glukosa yang terlalu tinggi pada medium dapat menurunkan produktivitas jamur untuk menghasilkan enzim ekstraseluler.
Hasil pada Gambar 2 menunjukan aktivitas lakase yang tinggi. Telah diketahui bahwa lakase dari banyak spesies jamur menjadi kelompok isoenzim yang dikodekan oleh gen dari family jamur tersebut, misalnya, Agaricus bisporus, Cerrena unicolor, Trametes sanguinea, dan Trametes versicolor (Wang dkk., 2018). Penggunaan medium kirk dalam penelitian ini tanpa penambahan sumber lignin, sehingga enzim Lip dan Mnp yang terukur tidak menunjukan hasil yang baik.
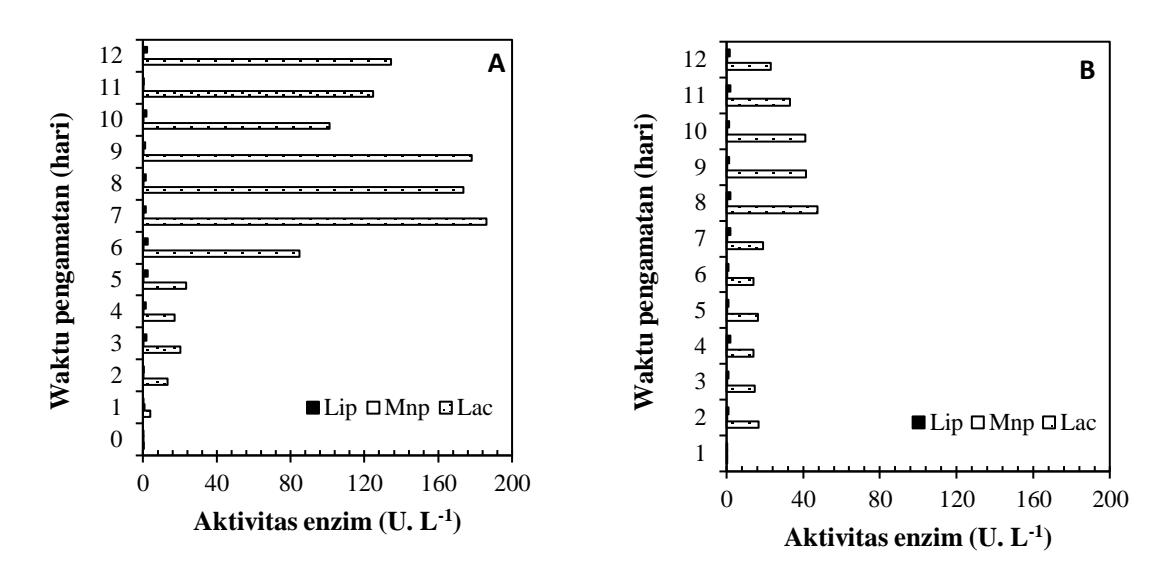
Gambar 2. Grafik aktivitas enzimatis (A) Trametes versicolor; (B) Pleurotus ostreatus
Pada penelitian terdahulu, menunjukkan bahwa senyawa fenolik dan nonfenolat teroksidasi oleh senyawa yang berasal dari reaksi Fenton yang dimediasi oleh kompleks molekul rendah Fe3+, seperti MnP, dan laccase dapat mengoksidasi senyawa lignin non-fenolik dengan kehadiran Fe3+ atau Mn2+/Mn3+ tersebut (Munoz dkk., 1997). Oleh karena itu, medium Kirk yang digunakan sebagai substrat utama untuk pertumbuhan jamur pelapuk putih memiliki komposisi ion-ion logam yang dibutuhkan untuk menjadi mediator. Sehingga, radikal hidroksil yang dihasilkan melalui reaksi berbasis Fenton non-enzimatik tersebut memulai oksidasi substruktur non-fenolik, dan sebagian senyawa non-fenolik yang teroksidasi kemudian dapat
dioksidasi lebih lanjut oleh enzim peroksidase dan/atau lakase (Arantes & Milagres, 2006). Dari hasil penelitian ini, dapat diamati bahwa pada kedua isolat, aktivitas enzim lakase lebih tinggi dibandingkan Mnp dan Lip. Sehingga, untuk percobaan penyisihan warna dengan SFF hanya aktivitas lakase yang akan diamati sebagai ekstraseluler enzim yang dominan diproduksi dan isolate T. versicolor menghasilkan enzim lakase dengan aktivitas yang lebih tinggi dibandingkan isolat P. ostreatus.
Biomasa kering Trametes versicolor dan pH pada SFF
Hasil inkubasi selama 5 hari kultur T. versicolor yang diujikan dengan pewarna reaktif berdasarkan biomasa kering sel pada medium SFF dapat diamati pada Gambar 3A Pada akhir pengamatan penyisihan warna RB5 menunjukan hasil biomassa kering tertinggi, yaitu 4,66 g/L, kemudian RB4 dan RO16 (4,3 g/L), serta RR120 (4,28 g/L). Biomasa kering pada masingmasing penambahan warna meningkat dari hari ke-1 sampai hari ke-5. Peningkatan pertumbuhan biomasa jamur dalam reaktor SFF menunjukan kemampuan toleransi jamur terhadap pewarna tekstil yang digunakan.
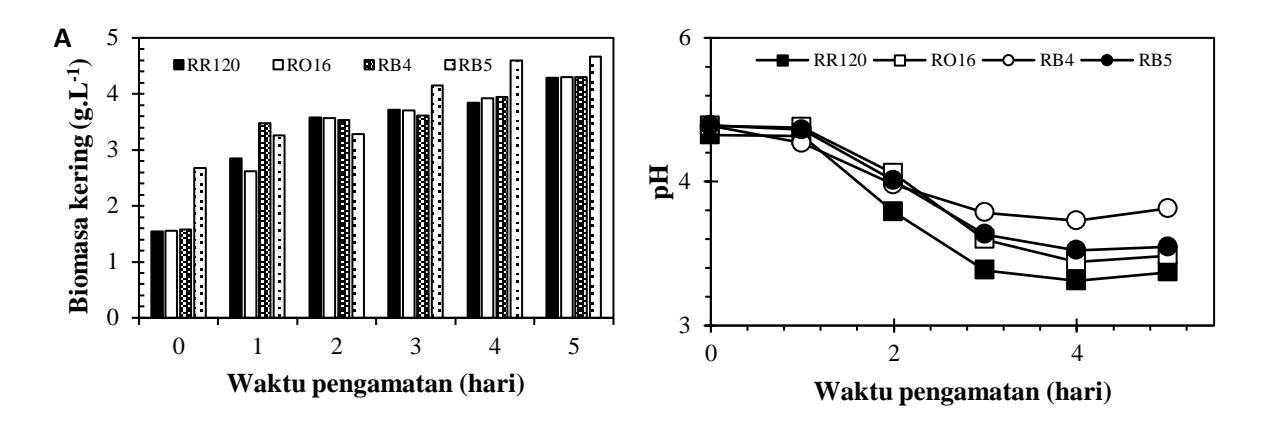
Gambar 3. Grafik biomasa kering dan pH pda SFF
Pada pengamatan pH yang terukur (Gambar 3B), penurunan pH menunjukan adanya aktivitas biologis yang menghasilkan asam organik seperti malat, oksalat, fumarat, dan glioksalat diproduksi selama fermentasi jamur pelapuk putih. Asam-asam ini bertanggung jawab atas penurunan pH. Secara alamiah jamur memiliki sistem buffer untuk tetap menjaga kualitas enzim yang digunakan. Hal tersebut tentunya menguntungkan karena enzim dapat bekerja pada rentang pH yang luas.
Percobaan Penyisihan warna oleh Trametes versicolor pada SFF
Laju penyisihan pewarna azo pada awal sampai akhir pengamatan dapat dilihat pada Gambar 4A selama 5 hari masa inkubasi. Pada penambahan pewarna jenis antrakuinon RB4 persen penyisihan warna yaitu sebesar 99,99% disisihkan lebih baik dibandingkan jenis pewarna azo lainnya, RB5 (98,03 %), RR120 (90,56 %) serta RO16 (63,52 %). Pada percobaan ini enzim lakase aktivitasnya terus naik selama waktu inkubasi, aktivitas enzim yang terukur secara berturut-turut pada akhir perlakuan untuk RB4, RB5, RR120 dan RO16 adalah 547 U.l-1 , 382 U.l-1 , 325 U.l-1 dan 294 U.l-1 (Gambar 4B).
Besarnya aktivitas enzim lakase yang terukur sebanding dengan persen penyisihan warna pada percobaan ini. Hal tersebut dapat membuktikan bahwa mekanisme penyisihan warna tersebut dipengaruhi oleh aktivitas lakase. Dari penelitian ini, percobaan dengan penambahan pewarna RB4 yang merupakan jenis antrakuinon memiliki persen penyisihan yang lebih baik dibandingkan tiga jenis pewarna azo lainnya, pada konsentrasi pewarna 150 mg/L.
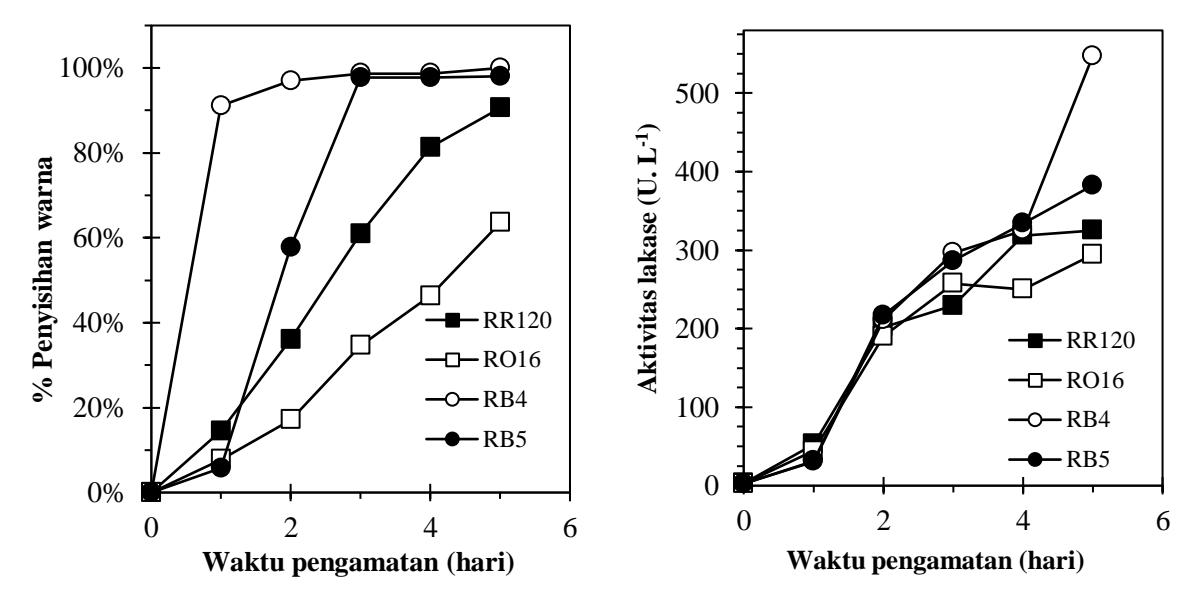
Gambar 4. Grafik (A) Penyisihan pewarna sintetik konsentasi 150 mg/L; (B) Aktivitas lakase pada masing-masing pewarna
Kemampuan T. versicolor atau lakasenya untuk penyisihan pewarna yang berbeda ditunjukkan dalam beberapa penelitian. T. versicolor sebelumnya telah menunjukan kemampuan untuk penyisihan sejumlah pewarna baik seluruhnya atau sebagian dalam kondisi di agitasi (Ramsay & Nguyen, 2002). Osma dkk., (2010) menunjukkan bahwa penyisihan warna RBBR oleh lakase yang diimobilisasikan memiliki penyisihan maksimum (44%)
diperoleh dalam 42 jam terutama karena peran aktivitas enzim, produk transformasi RBBR yang terbentuk oleh pengolahan dengan enzim lakase menunjukkan lebih sedikit fitotoksisitas daripada pewarna induknya. Menurut hasil percobaan penyisihan warna dalam SFF, penulis menyimpulkan bahwa enzim ekstraseluler dari organisme ini dapat secara efisien menghilangkan berbagai jenis pewarna tekstil.
Hasil tersebut menekankan peran lakase dalam mendegradasi pewarna antrakuinon. Seperti hasil studi mengenai jalur transformasi RBBR oleh imobilisai lakase yang diusulkan oleh Osma dkk., (2010). Hubungan antara efisiensi penyisihan warna dan aktivitas enzim dari jamur pelapuk putih telah dilaporkan sebelumnya (Banat dkk., 1996). Dalam literatur, ditekankan bahwa untuk penyisihan yang lebih efisien, berbagai faktor seperti optimalisasi bahan-bahan medium utama, pengamatan pertumbuhan jamur, peningkatan aktivitas enzim, investigasi laju penyisihan warna harus dipertimbangkan.
Scanning Perubahan Panjang gelombang pada pewarna reaktif setelah perlakuan
Selain dengan pengukuran konsentrasi, dilakukan juga scanning panjang gelombang pewarna teroksidasi. Laju penyisihan pewarna teroksidasi menunjukkan bahwa terjadi penurunan nlai absorbansi pada Panjang gelombang sinar tampak untuk keempat pewarna. Hasil scanning panjang gelombang pewarna ditunjukkan oleh Gambar 5.
Pada pewarna tekstil RR120 puncak spektrum menurun pada rentang sinar tampak namun tidak bertambah pada rentang spektrum serapan elektronik dari protein terutama ditandai pada daerah ultraviolet (185-320 nm). Hal tersebut dapat mengindikasikan tidak terbentuknya hasil samping produk hasil penyisihan warna. Berbeda halnya pada pewarna tekstil RB4, RB5 dan RO16, pada spektrum ultraviolet (185-320 nm), adanya penambahan absorbansi puncak spektrum yang dapat mengindikasikan terjadinya biotransformasi pewarna menjadi hasil samping produk lainnya.
Konsentrasi pewarna yang digunakan juga mempengaruhi kerja enzim dalam menyisihkan pewarna tekstil. Beberapa penelitian menunjukkan bahwa kemampuan enzim dalam menyisihkan warna memiliki kemampuan berbeda tergantung konsentrasi pewarna yang digunakan (Ratanapongleka & Phetsom, 2014). Penelitian tersebut menggunakan konsentrasi pewarna dari konsentrasi 20 mg/l hingga 140mg/l.
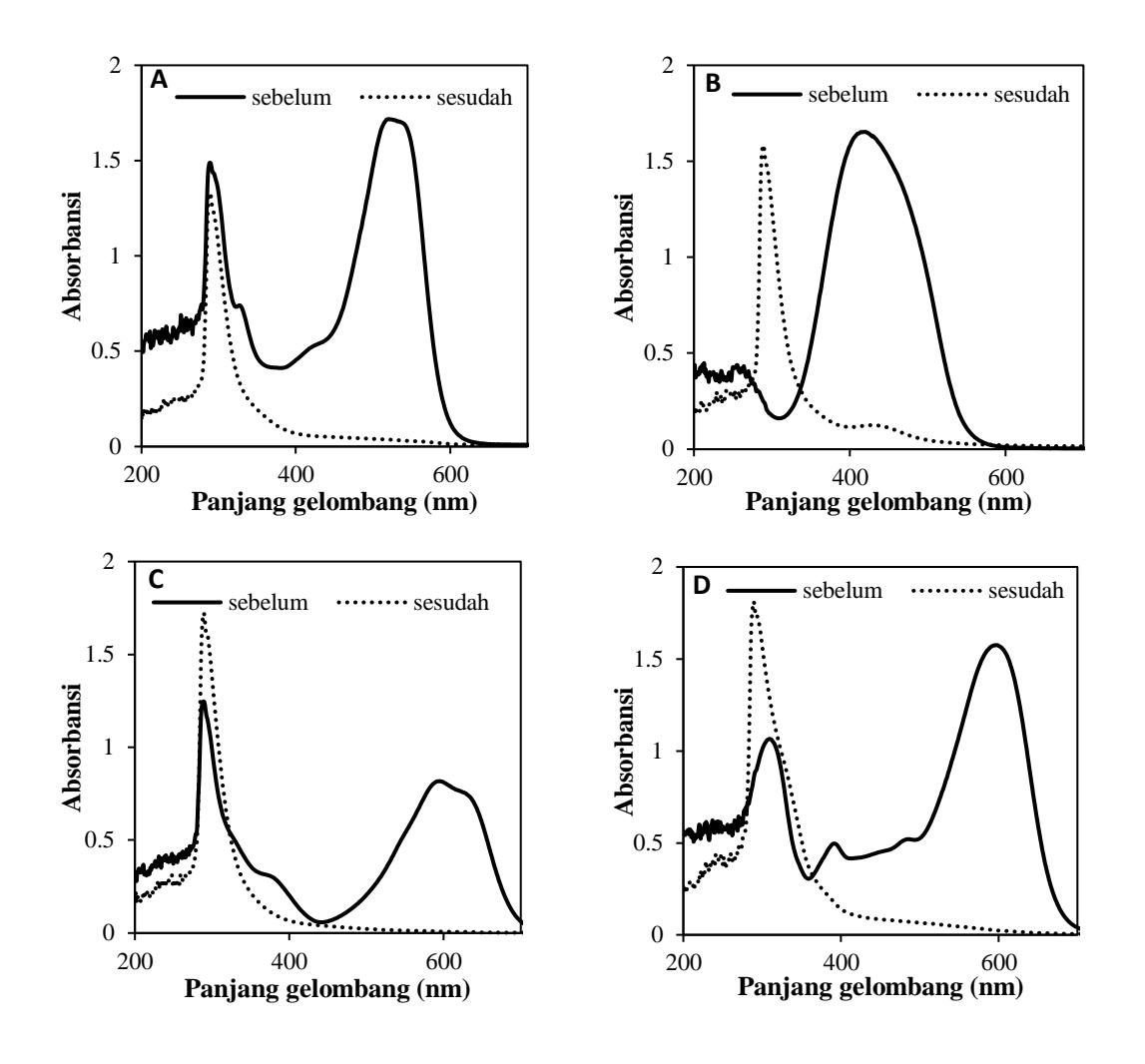
Gambar 5. Scanning Perubahan Panjang gelombang pada pewarna reaktif sebelum dan sesudah perlakuan (A) RR120; (B) RO16; (C) RB4; dan (D) RB5
Hilangnya (total) puncak tunggal pada panjang gelombang maksimum (λmax) dari maing-masing pewarna menunjukkan bahwa proses tingkat penyisihan warna terlihat sampai pada titik dimana secara visual warna menjadi hilang. Pewarna terutama azo dikenal dengan penggunaan pada kuantitas kecil karena warna dan kecemerlangannya yang khas. Bentuk murni pewarna biasanya direpresentasikan oleh warna asli dan panjang gelombang pada absorbansi tertinggi (λmax) ditentukan dengan pemindaian UV. Hasilnya (UV-Vis spectra), disintegrasi puncak yang diamati pada λmax setelah penyisihan pewarna terbukti karena penurunan absorbansi di wilayah UV.
Uji stabilitas pH pada ekstrak kasar enzim lakase dari T. versicolor
PH media kultur merupakan faktor penting dalam produksi lakase dan dapat mengganggu aktivitas enzim dan stabilitasnya (Sharma dkk., 2013). pH optimal ekstrak kasar
enzim lakase terhadap substrat ABTS yaitu pada pH 2,4. Sedangkan aktivitas relative enzim yang diamati pada pH 2,2 buffer asam sitrat-Na2HPO4 atau buffer McIlvaine (93,8%). Lebih dari 50% aktivitas enzim berkurang pada pH 3,8 dan lebih dari 95% aktivitas berkurang pada pH 5,8. Kemampuan oksidasi enzim hampir berkurang sepenuhnya pada kisaran pH lebih dari 6,2. PH optimal untuk aktivitas lakase umumnya rendah, berkisar antara 3,0 dan 5,0 (More dkk., 2011). Pada hasil penelitian ini pH optimal yang terbaik berkisar antara 2,2 sampai 4,4 dan kemudian aktivitas enzim terus menurun dan tidak terdeteksi adanya aktivitas dari rentang pH 6,8 sampai 8,0. PH optimal untuk aktivitas lakase tergantung pada substrat yang dipilih (Patel dkk., 2014), pH optimum dapat berubah menjadi kurang dari 5,0 untuk ABTS sedangkan untuk guaiacol agak lebih tinggi (pH 5,0-7,0) untuk MPD (2,6-Dimethoxyphenol) dan syringaldazine (Sharma dkk., 2013). Hal tersebut dapat dikaitkan dengan mekanisme reaksi lakase, yang melibatkan abstraksi elektron atau hidrogen yang akan dipengaruhi oleh perubahan pH. Selain itu, reaksi oksidasi tergantung pada redoks potensial substrat. pH mempengaruhi keadaan ionisasi substrat dan karenanya memengaruhi kapasitasnya untuk bertindak sebagai substrat pereduksi (Majcherczyk dkk., 1999).
Suhu optimal untuk ekstrak kasar enzim lakase T. versicolor adalah 20 0C. Sekitar 67% dari total aktivitas relatif tetap terukur ketika diuji hingga suhu 50 0C. Lebih dari 91% total aktivitas berkurang setelah 60 menit inkubasi pada suhu 55 0C. ekstrak kasar enzim lakase juga memanifestasikan stabilitas yang sangat tinggi dalam kisaran suhu uji 20-50 0C selama inkubasi 60 menit. Setelah diinkubasi pada 20-50 0C selama 60 menit, sebagian besar aktivitas pengoksidasi enzim tetap ada. Ketika diinkubasi pada suhu diatas 550C, enzim mengalami penurunan aktivitas pengoksidasi dan hampir tidak terukur pada suhu 70-80 0C.
Percobaan penyisihan warna menggunakan ekstrak kasar enzim lakase
Dalam Tabel 1, hasil penyisihan RR120, RO16, RB4 dan RB5 oleh ekstrak enzim kasar dengan aktivitas spesifik enzim sebesar 1842,91 U.L-1 disajikan. Pada penyisihan RB4 dengan waktu kontak 60 menit menunjukkan persentase penyisihan di atas 90%. Pewarna seperti pewarna antrakuinon adalah substrat yang baik untuk lakase dan penyisihan warna yang terjadi sebanding dengan aktivitas enzim (Wong & Yu, 1999).
Tabel 1. Penyisihan warna oleh ekstrak kasar enzim lakase T. versicolor
| Strain | Pewarna | Tipe penyisihan warna/ Medium | pH/Suhu/ Agitasi/ waktu kontak | Penyisihan warna (%) | Ref. |
|---|---|---|---|---|---|
| Trametes | RBBR (50 mg/l) | Medium terfiltrat/ | 30 °C pH 4,0 | 85,2% | |
| trogii | RB 4 (35 mg/l) | solid soybean cake | kondisi statis | 69,6% | Zheng |
| SYBC- | AB 129 (83,3 mg/l) | dan wood shavings | 30 menit | 45,6% | dkk, |
| LZ | AR 1 (10 mg/l) | substrate | 90,2% | 2011 | |
| RB5 (18,3 mg/l) | 65,4% | ||||
| Trametes | RR120 (150 mg/l) | Medium terfiltrat/ | 20 °C pH 2,4 | 6,61% | |
| versicolor | RO16 (150 mg/l) | medium Kirk | Agitasi 140 rpm | 4,95% | Studi |
| RB4 (150 mg/l) | 60 menit | 99,84% | ini | ||
| RB5 (150 mg/l) | 0,56% |
Penggunaan kultur jamur utuh untuk penyisihan warna tidak mudah diterapkan dalam skala besar. Selain itu, penggunaan enzim yang dimurnikan atau diimobilisasi akan meningkatkan biaya proses penyisihan warna (Levin dkk, 2010). Filtrat kultur kasar enzim memiliki beberapa keunggulan. Proses produksinya tidak mahal, mereka mungkin termasuk mediator lakase alami yang disekresikan oleh jamur (Johannes & Majcherczyk, 2005), dan di samping itu, faktor lain dalam medium seperti protein, sisa makronutrien, dan metabolit ekstraseluler yang larut dapat menstabilkan enzim kasar. Dalam hal ini penghilangan warna oleh filtrat kultur kasar enzim dapat diselesaikan secara efisien dalam 60 menit pada RB4, yang jauh lebih pendek daripada proses penyisihan warna oleh kultur jamur pada SFF (sekitar 5 hari). Selain itu, penyisihan pewarna menggunakan enzim kasar dapat menghindari penghambatan pewarna pada pertumbuhan jamur. Dengan demikian penyisihan warna menggunakan enzim kasar lebih menguntungkan. Berikut adalah stuktur kimia dari pewarna RB4 yang aktif didegradasi oleh ekstrak kasar enzim lakase (Gambar 6).
Gambar 6. Struktur kimia pewarna tekstil RB4
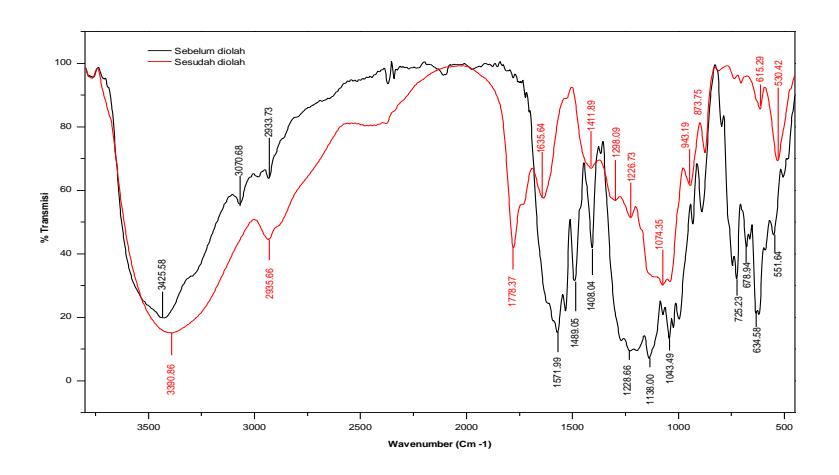
Gambar 7. Hasil analisis FTIR pewarna RB4 sebelum dan sesudah diolah
Puncak pada 3425 cm<sup>-1</sup> menunjukkan adanya ikatan -N-H- dalam pewarna asli, Amina primer =N—H dan amina sekunder -NH2 sering ada dalam struktur pewarna. Rentang nilai absorpsi dari peregangan N-H terkadang sulit dibedakan dengan ikatan hidrogen O-H. Penyerapan (% transmisi) N-H biasanya lebih tajam karena kecenderungan yang jauh lebih lemah dari kelompok N-H untuk membentuk ikatan hidrogen (Manfred dkk., 1997). Namun, dalam kasus pewarna reaktif, baik kelompok O-H dan N-H kemungkinan ada dalam struktur pewarna.
Sementara peregangan gugus -C-H pewarna teramati pada 2933-3070 cm<sup>-1</sup>, yang menghilang dalam spektrum metabolit (hasil pengolahan). Selain itu, pemutusan cincin antrakuinon juga dibuktikan oleh puncak yang semakin lemah di dekat 1571 cm<sup>-1</sup>, yang bertanggung jawab untuk kombinasi getaran peregangan C=O terkonjugasi dengan C=C (Miao dan Tao, 2008). Secara khusus, ikatan penghubung antara cincin antraquinone dan cincin mono benzene dengan cepat terputus karena menurunnya intensitas puncak pada 1269 cm<sup>-1</sup> untuk kelompok amina aromatik.
Gugus karbonil dalam kromofor pada umumnya cenderung dikonjugasikan dengan ikatan rangkap atau cincin aromatik (sering pada kedua substituen pada karbonil) (Miller, 1977). Kelompok C=O dapat tumpang tindih dengan bending N-H dan C=C terkonjugasi dengan cincin aromatik dalam spektrum IR. Oleh karena itu, tidak dapat digunakan untuk mengidentifikasi apakah pewarna tersebut termasuk pewarna reaktif jenis antrakuinon. Keton, =C=O, dapat ditampilkan dalam struktur reaktif biru. Kelompok C=N dapat ditemukan dalam beberapa pewarna reaktif. Hal tersebut mungkin tumpang tindih dengan senyawa karbonil dan N-H bending dalam spektrum IR. Dengan mengacu pada pewarna reaktif, C=N sering terdapat dalam sistem siklik terkonjugasi.
Kehadiran puncak pada 1269 cm-1 merupakan amina aromatik yang ditemukan dapat diabaikan setelah penyisihan pewarna karena puncaknya yang menghilang. Pada struktur cincin aromatic RB4 dengan puncak bilangan gelombang antara 619 dan 725 cm-1 menghilang setelah proses pengolahan, yang menunjukan aktivitas biotransformasi oleh ekstrak kasar enzim lakase.
Cincin aromatik dapat diidentifikasi oleh beberapa daerah band di spektrum IR. Garam sulfonat, gugus R-SO3- adalah komponen penting dari pewarna reaktif serta senyawa kloroaromatika (C-Cl) sering pula terdapat dalam struktur pewarna reaktif. Senyawa aromtaik ada dalam struktur warna cyan, magenta, dan hitam dalam penyerapan yang lemah, tetapi intens dan luas dalam pada pewarna kuning.
Setelah pengolahan menggunakan ekstrak kasar enzim, spektrum RB4 menunjukkan hilangnya berbagai puncak. Pada saat yang sama, muncul puncak baru di 1298 cm-1 dan 1778 cm-1 masing-masing menggambarkan adanya getaran CO asam dan deformasi C = O asam serta aldehida yang dapat disebabkan oleh produk sampingan yang terbentuk selama proses penyisihan RB4. Hasilnya menentukan bahwa ekstrak kasar enzim lakase T. versicolor secara efektif menghilangkan senyawa terhalogenasi serta gugus amina dari RB4 seperti yang dijelaskan sebelumnya yaitu penyisihan warna oleh Proteus mirabilis dan Proteus vulgaris serta cyanobacteria (Parmar & Shukla, 2015; Afreen dkk, 2017).
KESIMPULAN
Jamur pelapuk putih jenis Pleurotus ostreatus, Trametes versicolor, Marasmiellus palmivorus, dan Ganoderma sp. memiliki potensi yang berbeda dalam mendegradasi pewarna tekstil reaktif. Secara in vitro pada medium PDA spesies P. ostreatus dan T. versicolor aktif menyisihkan keempat pewarna yang digunakan, yaitu antrakuinon (reactive blue 4), monoazo (reactive orange 16) dan diazo (reactive red 120 dan reactive black 5). Evaluasi aktivitas enzim kedua spesies menunjukan bahwa lakase merupakan enzim yang dominan terukur pada medium nutrient yang digunakan, lebih lanjut T. versicolor memiliki aktivitas lakase terbaik, serta mampu menyisihkan pewarna reaktif sebesar 60-90% pada SFF. Aktivitas lakase dapat bekerja pada rentang pH 2,2 hingga 4,4, serta pada suhu 20–50 0C. 5. Percobaan penyisihan warna dengan ekstrak kasar enzim lakase terbaik yaitu pada RB4 sebesar 99,84%. Hasil studi toksisitas menggunakan Bacillus sp. terhadap pewarna RB4 sebelum dan sesudah diolah, menunjukan hasil metabolitnya telah berkurang toksisitasnya dibandingkan senyawa induknya.