
1. Introduction
Leathers have long been used as a raw material for various items, such as clothing, furniture, and accessories, owing to their tough yet flexible nature and thin structure. The global leather goods market size is estimated to grow at a combined annual growth rate of 9.5% [1]. The primary sources of this leather material come from livestock such as cows, sheep, and horses, which raise environmental concerns about deforestation for pastureland, low efficiency, and lengthy production processes [2]. Therefore, there is a growing need for an alternative material that can be produced on a large, controlled scale, be environmentally friendly and efficient, and possess characteristics akin to genuine leather. These criteria align with mycelium that grows rapidly, efficiently utilizes nutrients, and can be processed into materials with properties similar to genuine leather. One potentially suitable species is Ganoderma lucidum, recognized and used in several vegan leather industries for its physical resilience, color stability, and equal or better flexibility compared to cowhide and polyurethane (PU) [3]. However, further research is needed to determine the optimal medium for producing G. lucidum spawn as an industrial-scale biomaterial feedstock. [2].
Ganoderma lucidum thrives well in common mushroom media, such as PDA, which provides readily available carbon and nitrogen sources. As a member of the white-rot fungus
group, G. lucidum can also utilize other plant-derived carbon sources, such as straw, wood, and cereals [4,5]. This ability to degrade lignocellulose allows it to grow on substrates, activating the production of cellulose-degrading enzymes like cellobiohydrolase, β-glucosidase, and lytic polysaccharide monooxygenase, alongside lignin-modifying enzymes like lignin peroxidase, manganese peroxidase, and laccase, which conduct oxidative degradation and alkyl-aryl bond cleavage in phenolic groups using peroxides and Mn2+ ions [6]. This fungus is commonly found growing on wood plants and, in some cases, non-wood plants, acting as a parasite that leads to tree death and decay. Typically, the wood used for mushroom cultivation, including G. lucidum, is Albizia wood (sengon) in sawdust form due to its high availability and low cost. Sengon sawdust is rich in cellulose, hemicellulose, and lignin which are the main carbon sources for mushroom growth. However, alternative sources are now needed due to increasing demand and rising costs. Potential alternatives can be found in bamboo [7], and oil palm as one of G. lucidum natural habitats [8]. Additionally, grain additions, such as corn, are commonly used as supplements [4].
In Indonesia, bamboo is a widespread plant utilized in various regions for diverse purposes. The total processed bamboo production in Indonesia reached 8.65 million stems/ year in 2014. The advantage of using bamboo powder as a mushroom-growing medium lies in its high carbon supply as a nutrient source, with a cellulose content of 42.4–53.6% and a lignin content of 19.8–26.6% [9]. The high protein, phosphorus, and potassium content can also accelerate fungal growth [10].
The second substrate in this study is oil palm (Elaeis guineensis), one of Indonesia's largest productivity commodities. Oil palm producers typically generate substantial quantities of empty palm oil fruit bunches (EFB), which are commonly discarded as waste. Nutritionally, EFB contains about 23.7–65.0% cellulose, 20.58–33.52% hemicellulose, and 14.1–30.45% lignin. Their high cellulose content makes EFB a potential raw material for mushroom cultivation substrates [11].
Corn is often selected as a growth medium due to its rich nutrient content, which supports mushroom mycelium growth, including carbohydrates at 69.0%, protein at 12%, fat at 6.0%, fiber at 2.2%, and other organic matter at 15% [12]. The addition of corn aims to expedite nutrient absorption into mycelium cell walls. This is because corn's simpler structure and lower lignin content compared to wood make its powder molecules more readily absorbed by extracellular mycelial enzymes [13]. The effectiveness of corn in optimizing mushroom growth can be enhanced by a pre-processing method that dries and transforms it into cracked corn, which yields better results than undried corn and other cereal types [14].
This research aims to determine the influence of adding bamboo sawdust and EFB on the growth rate of G. lucidum in the medium, identify the best concentrations of bamboo sawdust and EFB for F1 G. lucidum spawn production, and examine the effect of modifying solid medium substrates using bamboo sawdust or EFB (on growth rate & LOM (loss of organic matter) in mycelium growth compared to cerealsengon mediums.
2. Methodology
This research was conducted at the Technology Laboratory XI, Faculty of Life Sciences and Technology, Bandung Institute of Technology (ITB), and at the Mycotech Eco Factory Laboratory from October 2022 to May 2023. The G. lucidum isolates were obtained from the Microbiology Laboratory, Center for Bioscience and Biotechnology Research, ITB. Bamboo sawdust and sengon sawdust were obtained from The Wood Laboratory, Jatinangor Campus, ITB, and EFB was obtained from a palm oil plantation in Parung, Bogor, West Java.
2.1 Preparation of isolates and media
The G. lucidum isolates used for the treatment were cultivated on PDA (Potato Dextrose Agar) medium in a petri dish and PDB (Potato Dextrose Broth) medium in an Erlenmeyer flask. Bamboo sawdust, EFB, and sengon sawdust were ground using a grinding machine. In this study, three stages were used to obtain the optimal growth medium for G. lucidum.
2.2 Experimental Design
This study employs a completely randomized design (CRD) with two treatments at various concentrations: bamboo sawdust and EFB (stage 1 and stage 2), and in stage 3, 3 treatments: bamboo sawdust, EFB, and sengon sawdust as a control. In the first stage, six variations of bamboo sawdust and EFB concentrations were added to PDA media. The second stage used five variations of bamboo sawdust and EFB concentrations added to PDB media, while the third stage used three treatment variations, namely a mixture of cracked corn and bamboo sawdust, a mixture of cracked corn and EFB, and a mixture of cracked corn and sengon sawdust as a control, where each treatment had three replications.
2.3 Stages of the experiment
In the first stage, G. lucidum was inoculated into two media, namely PDA with added bamboo sawdust and PDA with added EFB, each at concentrations of 0% (PDA
100 DOI: 10.5614/3bio.2026.8.2.5
medium only), 2%, 4%, 6%, 8%, and 10% (w/v). Incubation occurred at 25°C for eight days until the mycelium covered the entire surface of the petri dish medium. Radial mycelium measurements were taken every 24 hours. In the second stage, G. lucidum was inoculated into two types of media, namely PDB media supplemented with bamboo sawdust and PDB media supplemented with EFB, each with a concentration of 0% (PDB media only), 25%, 50%, 75%, and 100% (w/v). The control treatment was prepared with a composition of 100% PDB and 0% bamboo sawdust or EFB. Agar medium was added separately to each variation at 1.5% (w/v), and distilled water was added to adjust the moisture content to 65%. The medium was sterilized in an autoclave at 121°C and 1 atm for 15 minutes. In the third stage, the treatment to optimize G. lucidum growth on bamboo sawdust and EFB media was carried out by modifying the growth media. The control composition consisted of 50% cracked corn (weight/ weight) and 50% sengon sawdust (w/w). The modified media optimization treatments using bamboo sawdust and EFB were as follows: modification 1 consisted of bamboo sawdust with added cracked corn, and modification 2 consisted of cracked corn with added EFB in a 1:1 ratio. Distilled water was added to adjust the moisture content to 65%. Incubation continued at 25°C for eight days until the mycelium covered the entire surface of the petri dish medium. Radial mycelium measurements were taken every 24 hours. Inoculation was also carried out in glass jars using the control medium of cornsengon, the modified bamboo, and the EFB media. Incubation lasted for 15 days at 25 °C until the mycelium covered the entire surface of the glass jar medium. Length measurements of downward mycelial growth were taken every 72 hours at six points.
2. 4 Research parameters
The first research parameter is the mycelium growth ratio, calculated using equation below:
\[X_t = \frac{(L_t - L_\theta)}{L_\theta} \quad [14]\]
The second research parameter is the mycelium growth rate, calculated using equation below:
\[Y_t = \frac{(L_t - L_{t-1})}{(T_t - T_{t-1})} \quad [15]\]
Mycelial growth was measured by dividing the Petri dish into 8 equal sections, each delimited by a line. At each observation time, the outermost visible portion of the mycelium was marked along the lines. The distance from the center to each mark was measured using a vernier caliper, obtaining radial data at eight points from the first to the eighth day.
The loss of organic matter was measured by collecting the medium containing mycelium from the glass jars and drying it gravimetrically [16] after 15 days. This involved weighing the sample before and after placing it in an 80°C oven for 24 hours. Post-drying weighing was promptly performed to prevent weight gain from adhering moisture. Calculations were conducted following the equation below:
\[\Delta OM = \frac{W_t - W_\theta}{W_\theta} \times 100\% \quad [17].\]
2.5 Data Analysis
Radial calculation data from stages 1 and 2 were analyzed using a Single-Factor ANOVA with a 95% confidence interval, using the Data Analysis feature in Microsoft Excel. Results indicating significant differences were further subjected to Duncan's Multiple Range Test (DMRT) for post-hoc analysis and annotation. For the results from stage 3, which involved two variations, the analysis was conducted using a t-test.
3. Results and discussion
The results were obtained by measuring mycelial diameter from day 0 to day 8 and calculating the ratio of mycelial diameter to the initial inoculum diameter. In stage 1, G. lucidum mycelium in media supplemented with bamboo sawdust grew faster (Fig. 1), as indicated by a higher radial ratio, especially at a bamboo concentration of 6%, which was significantly higher (p<0.05) than the other variations. This indicates that the additional nutrients from bamboo powder are utilized by G. lucidum, accelerating its growth by increasing organic content, such as protein and vitamin B, and inorganic substances, such as iron and calcium [7]. At higher bamboo concentrations, mycelium growth was not as optimal as at 6% due to the fungus requiring more time to adapt to higher concentrations of complex substrates [18]. Variations with EFB also showed increased growth rates, with the best variation at 10% concentration, indicating that G. lucidum could utilize EFB at relatively higher concentrations, likely because G. lucidum is known to grow on oil palm plants as a decay parasite, especially at the base of oil palm trunks [19]. Additionally, the carbon-to-nitrogen ratio in EFB is higher than in bamboo, and the higher its concentration in the medium, the more optimal the C:N ratio is for triggering G. lucidum growth [20].
Subsequently, cultures from the best-performing variations in stage 1 (6% Bamboo and 10% EFB) were transferred to the second-stage experiment, which involved higher concentrations of bamboo or EFB powder. The control showed better growth than the variations with added bamboo sawdust, except at the 25% concentration, as well as EFB (Fig. 2A,C). This occurred due to a significant decrease in dextrose levels concurrent with the decrease in the amount of
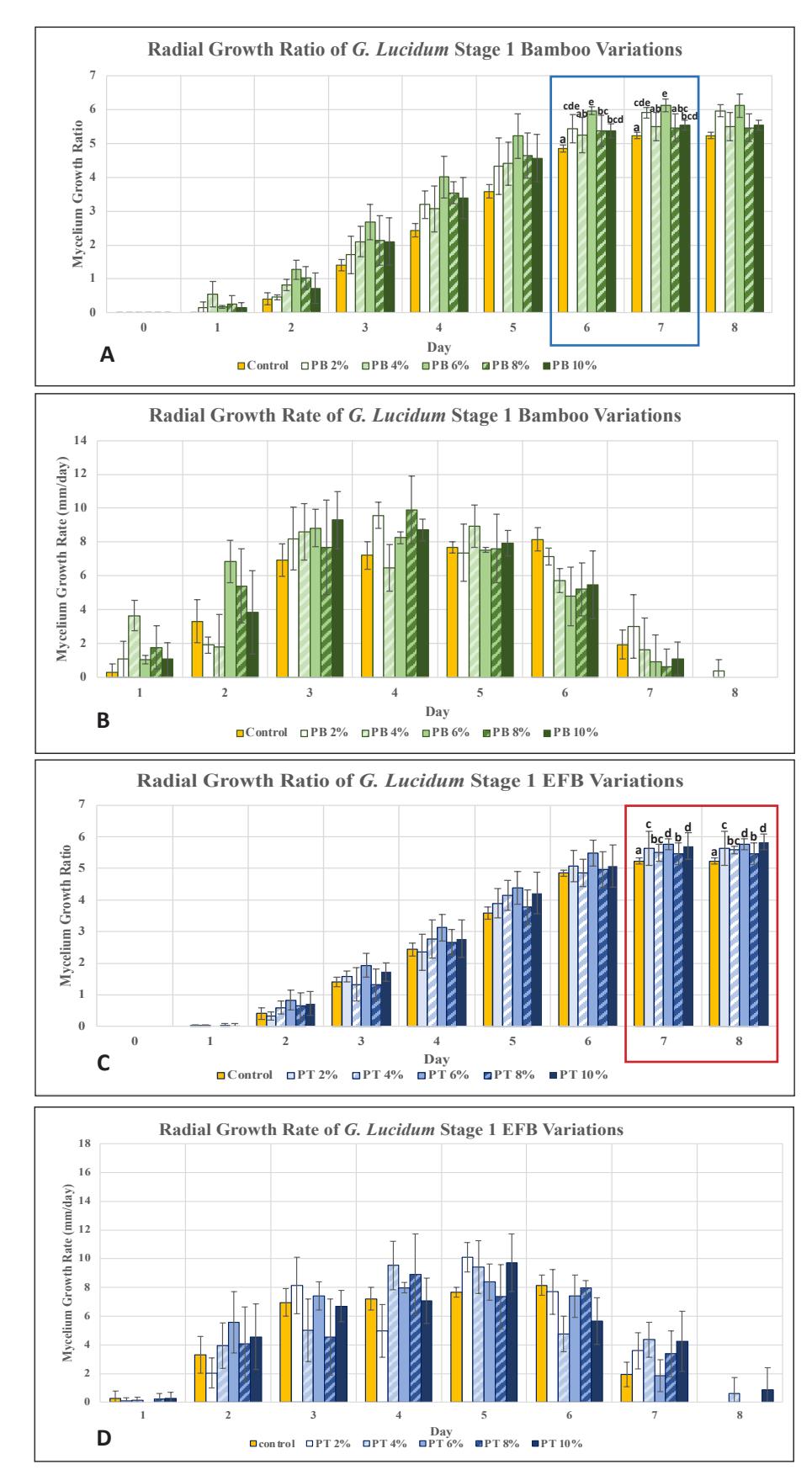
Figure 1. G. lucidum growth chart in stage 1: A. Ratio on PDA medium supplemented with bamboo; B. Rate on PDA medium supplemented with bamboo; C. Ratio on PDA medium supplemented with EFB; D. Rate on PDA medium supplemented with EFB
PDA used. Dextrose is a monosaccharide, the simplest form of sugar that can be readily absorbed by cells to produce energy and facilitate growth [21]. Meanwhile, the carbon sources in bamboo and EFB are lignocellulose, a polysaccharide that takes time to produce degradative enzymes to break glycosidic bonds [22,23], such as endo-\(\beta\)-1,4-glucanase, cellobiohydrolase, \(\beta\)-glucosidase [24], lignin-modifying enzymes (LME) like laccase, manganese peroxidase, and lignin peroxidase [6, 25].
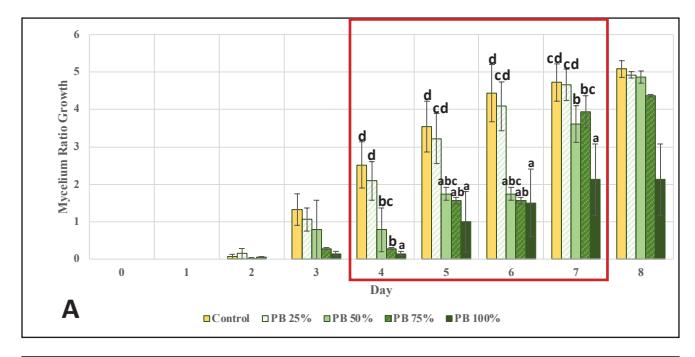
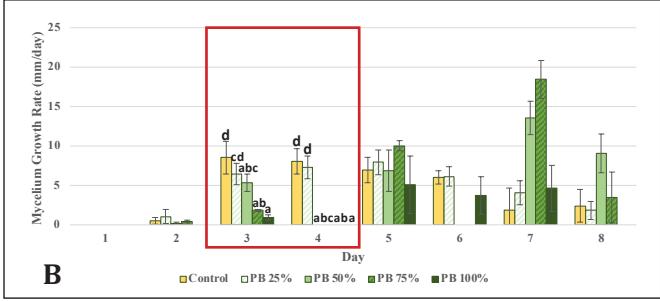
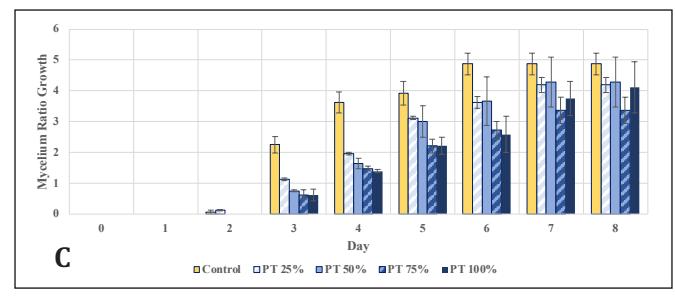
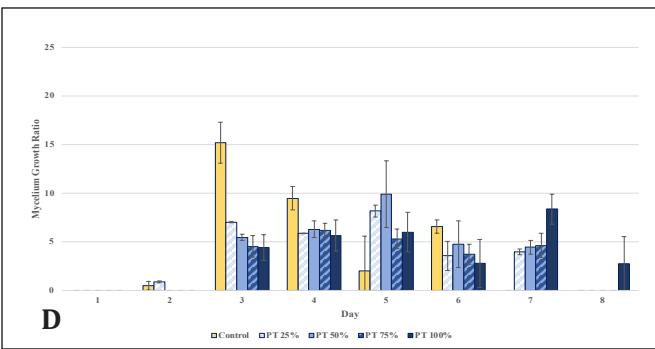
Figure 2. G.lucidum growth chart in stage 2: A. Ratio on PDA medium supplemented with bamboo; B. Rate on PDA medium supplemented with bamboo; C. Ratio on PDA medium supplemented with EFB; D. Rate on PDA medium supplemented with EFB
Upon observing the growth rate, the control rate peaked on the 3rd observation day and then declined through the 6th observation day. This might be due to the simplicity of dextrose, which can be directly utilized by G. lucidum; its concentration in the medium can decrease rapidly, limiting mycelial growth [26]. On the other hand, in EFB variations, the growth rate remained stable. It gradually increased due to the abundance of complex carbon sources, with concentrations significantly higher than in the previous stage. This leads G. lucidum to produce lignocellulose-degrading enzymes and to use wood substrates as a nutrient source while utilizing any remaining dextrose, resulting in relatively slow growth initially but continually increasing [6, 25]. The growth rate of the bamboo variations fluctuated. This could be attributed to the limitations of the radial method, which measures mycelial growth only horizontally and cannot assess thickness, vertical growth, or mycelial biomass [27]. In this stage, the best concentrations were 25% bamboo and 50% EFB, which vielded results equivalent to the control (p<0.05) in terms of growth ratio until the 8th day. Therefore, cultures at these concentrations were continued for inoculation into stage 3 medium, which uses only natural materials and no PDA.
In EFB, significant growth was observed earlier compared to the control, which emerged only after day 5, and bamboo after day 4, indicating better culture readiness with EFB, given the closer increase in powder concentration to the previous stage at 50%, compared to the bamboo variation at 25%, to degrade polysaccharides directly [25]. However, the growth thereafter stabilized and did not reach a ratio twice as high by day 8. These results need further confirmation by observing their vertical growth in jars.
The growth in the glass jar (Fig. 3) showed results similar to the radial growth observations. The culture on the control medium grew more slowly initially than on the modified medium. Subsequently, the growth ratio on the bamboomodified medium was significantly higher (p < 0.05) than the control on the ninth day of observation. Bamboo provides a dense source of carbon in the form of cellulose and is rich in other nutrients like protein, phosphorus, and potassium to support growth, which is relatively higher compared to most hardwoods, including sengon wood [10]. The better horizontal and vertical growth on the bamboo-modified medium demonstrated that bamboo powder can substitute sengon wood powder in F2 G. lucidum seed production. EFB variation showed rapid initial growth, but after 6 days there was no significant difference from the control until both reached maximum growth, filling the entire medium in the jar.
The produced biomass showed no significant difference between the control group, the modified bamboo group, and the modified EFB group (Fig. 4). There was also no significant difference in LOM (Fig. 5). When observing three-dimensional growth through radial and vertical growth in glass jars, the bamboo variety showed a faster growth rate
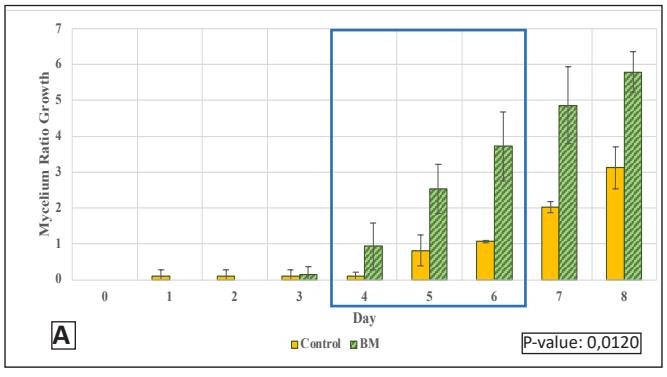
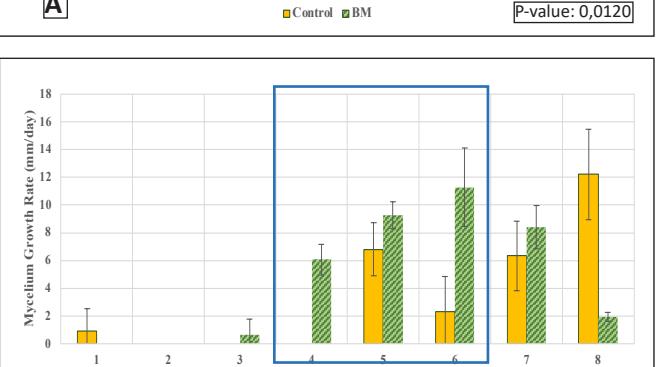
Day B Control BM P-value: 0,0083
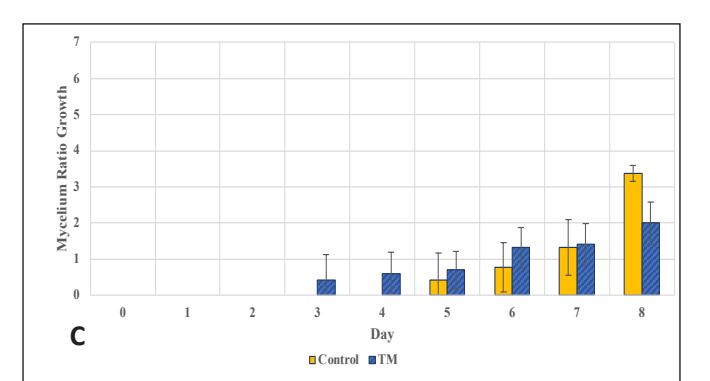
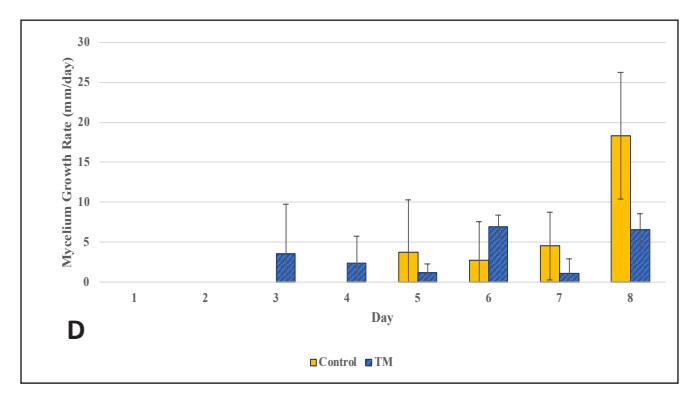
Figure 3. G.lucidum Growth Chart in Stage 3:
- A. Ratio in Bamboo-modified medium;
- B. Rate in Bamboo-modified medium; C. Ratio in EFBmodified medium;
- D. Rate in EFB-modified medium (Control = corn cracked-sengon, BM = corn-bamboo, TM = corn cracked-EFB)
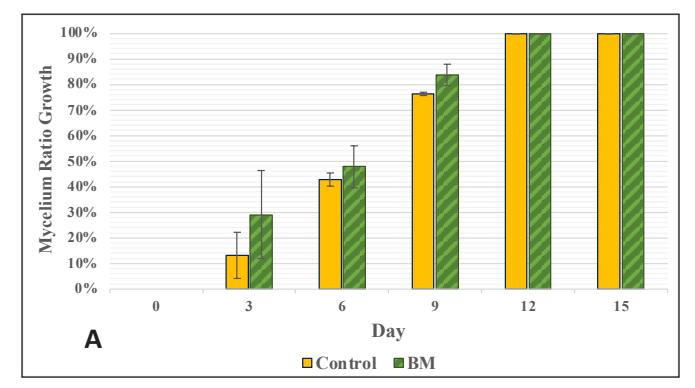
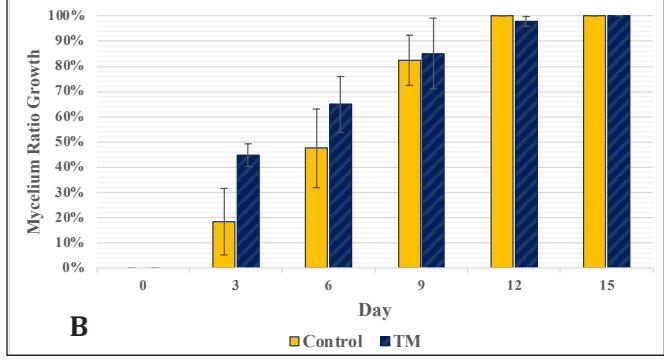
Figure 4. Chart of Mycelium Growth Percentage on Stage 3 Medium: A. Control and Bamboo sawdust Modification; B. Control and EFB Modification-EFB)
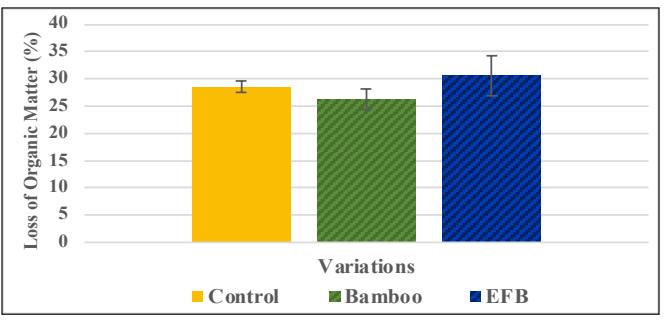
Figure 5. Graph of Comparison of Loss of Organic Matter (LOM) on modified medium: Sengon control, bamboo-modified, and EFB-modified
than the control group. However, the LOM figures did not differ significantly (p<0.05) from those of control group and the EFB variety. This suggests that using bamboo powder supports better expansion of G. lucidum but does not affect biomass production. This could be due to the softer particle texture of bamboo powder, resulting from its relatively low lignin content, and to its lower bulk density compared to typical hardwoods, which aids mycelium penetration into the substrate. However, this data may also be interpreted as rapid radial and vertical growth on bamboo media, indicating more efficient nutrient utilization by G. lucidum, resulting in higher growth rates without significant differences in LOM, as more nutrients are used for cell growth rather than converted into CO2. However, further research is needed to determine which possibility applies. One of the biggest challenges in the biomaterials industry is scaling processes from the laboratory to an economical industrial scale. Research on G. lucidum serves as an excellent model for studying and addressing this challenge.
4. Conclusion
The growth rate of G. lucidum on cereal medium modified with bamboo sawdust was faster than that on the control medium, but LOM did not show a significant difference. Meanwhile, the growth rate and LOM on the modified EFB medium did not differ significantly from those on the control medium. Bamboo sawdust and EFB can be considered alternative growth media for G. lucidum mycelium, as both support growth rates and LOM that are not significantly different from those in the control medium. To produce the best F1 culture, a 25% bamboo sawdust and 50% EFB mixture was used during the adaptation phase. Further studies will be needed, particularly related to stability testing by applying this formula on a larger scale (pilot project) to evaluate the consistency of results, production cost efficiency, and potential contamination, as well as testing several nitrogenrich supplements (such as rice bran or corn flour) in various concentrations added to the 25% bamboo and 50% EFB media to determine whether they affect increasing mycelium density and LOM values.